Screencap of Onion where some sort of scientist is announcing something in front of a Powerpoint slide of a clam
Last week, The Onion, a very serious journalistic publication, published a piece “Biologists Announce There Absolutely Nothing We Can Learn From Clams“. As a print subscriber I want to say I played a small part in this article, which I’ve actually hung on my office wall. But I want to take it a step further and write a line-by-line concurrence with everything they wrote!
WOODS HOLE, MA—Saying they saw no conceivable reason to bother with the bivalve mollusks, biologists at the Woods Hole Oceanographic Institution announced Thursday that there was absolutely nothing to be learned from clams.
Wow, I do know a researcher who studies clams at Woods Hole and actually love her work! Nina Whitney is now a prof at Western Washington University but until recently was a postdoc at WHOI studying how shells can serve as records of climate! I wonder who The Onion interviewed.
“Our studies have found that while some of their shells look pretty cool, clams really don’t have anything to teach us,” said the organization’s chief scientist, Francis Dawkins, clarifying that it wasn’t simply the case that researchers had already learned everything they could from clams, but rather that there had never been anything to learn from them and never would be.
Oh I don’t know a Francis Dawkins, but I’m sure they know their clams! It is true that their shells can look pretty cool. Bivalves include everything from Hysteroconcha dione, with its beautiful color and spines, to Tridacna gigas, which grows to 4.5 feet and weighs hundreds of pounds! And like an Onion, shells have growth layers, sometimes a new one every day, which someone could use to try to figure out how clams record what they eat and how the environment changes. But why would anyone do that?
Close-up view of a Hysteroconcha bivalve shell, showcasing its intricate ridges and coloration, and rows of long spines near the margin. Source
For me, I guess I haven’t learned anything from clams. I think I already knew in my heart that clams can live for >500 years. I already knew that mussels can filter several liters of water per hour, meaning that a colony of them can filter thousands of liters an hour. All this stuff is obvious, actually. Common sense.
“We certainly can’t teach them anything. It’s not like you can train them to run through a maze the way you would with mice. We’ve tried, and they pretty much just lie there.
It is ludicrous that clams could be taught anything or have anything approaching memory or thinking. It is only coincidence that scallops appear to clap their valves to swim, using their hundreds of eyes to navigate to a new location away from predators or toward food. It’s coincidence that they increase their feeding activity when shown a video of food particles. Some researchers have even claimed that giant clams can tell the difference between different shapes of objects! It is so dumb!
From what I’ve observed, they have a lot more in common with rocks than they do with us. They’re technically alive, I guess, if you want to call that living.
Also literally true! Their shell is a biomineral, in essence a living rock, made of calcium carbonate. They are alive in the sense they have a heart that beats, pushing hemolymph around their body. Their heart rate can increase or decrease with different stressors. Remember though, we always knew this. We didn’t learn it through something like science.
They open and close sometimes, but, I mean, so does a wallet. If you’ve used a wallet, you know more or less all there is to know about clams. Pretty boring.”
I myself have wasted time studying this. I attached sensors to giant clams to monitor their feeding activity. If I had learned anything, it might have been that they change their behavior between day and night, basking in the sun to help their photosynthetic algae in the day, and partially closing at night, with those behaviors changing based on how much chlorophyll is in the water. But remember! I didn’t learn it.
The finding follows a study conducted by marine biologists last summer that concluded clams don’t have much flavor, either, tasting pretty much the same as everything else on a fried seafood platter.
I can’t see how anyone would like to eat a bivalve. Especially not a fresh-caught scallop sauteed in butter or a plate of fried clams in New England. Never try that. Leave it to me!
Two giant clams near Eilat in the Northern Red Sea. To the left is the small giant clam, Tridacna maxima, and to the right is a mature individual of the rare endemic giant clam Tridacna squamosina, only found in the Northern Red Sea.
You are what you eat, and clams are too. We’re made of atoms, which come in “flavors” called isotopes, relating back to the mass of the atoms themselves (how many protons and neutrons they have). Nitrogen, for example, comes in two stable (non-radioactive) forms called nitrogen-14 and nitrogen-15. Much like scientists can track the composition of a person’s diet from the isotopes of their hair, researchers have used the isotopes of clams to figure out their diet.
Nitrogen isotopes provide us with a useful and detailed record of food webs. Plants and algae tend to have more of the light isotope of nitrogen in their tissues than the animals that eat them (primary consumers), and the animals that eat those animals have even higher nitrogen isotope values. We can measure the amount of “heavy” atoms of nitrogen with a unit called δ¹⁵N (“delta 15 N”). A carnivore at the top of the food chain will have a very high δ¹⁵N, while plants will be the lowest. Clams, typically being filter feeders, will usually have an intermediate value, since they’re eating a lot of phytoplankton (tiny microscopic floating algae) and zooplankton (animal plankton that eat other plankton).
But I study a special kind of clam, the giant clams, which have a cheat code enabling them to become giant: they have algae *inside* of their bodies. The algae make food using photosynthesis and share it with their hosts! In exchange, the clams provide the algae with a stable environment free of predators, plenty of fertilizer in the form of their own waste, and even channel extra light to help the symbionts grow faster. This partnership is called photosymbiosis, and is pretty rare in clams, though it is common in other animals like the corals that build the reefs where giant clams are found! Previous researchers have shown that giant clams have very low nitrogen isotopic values in their tissue, like a plant, because they get most of their nutrition from the algae, rather than filter feeding.
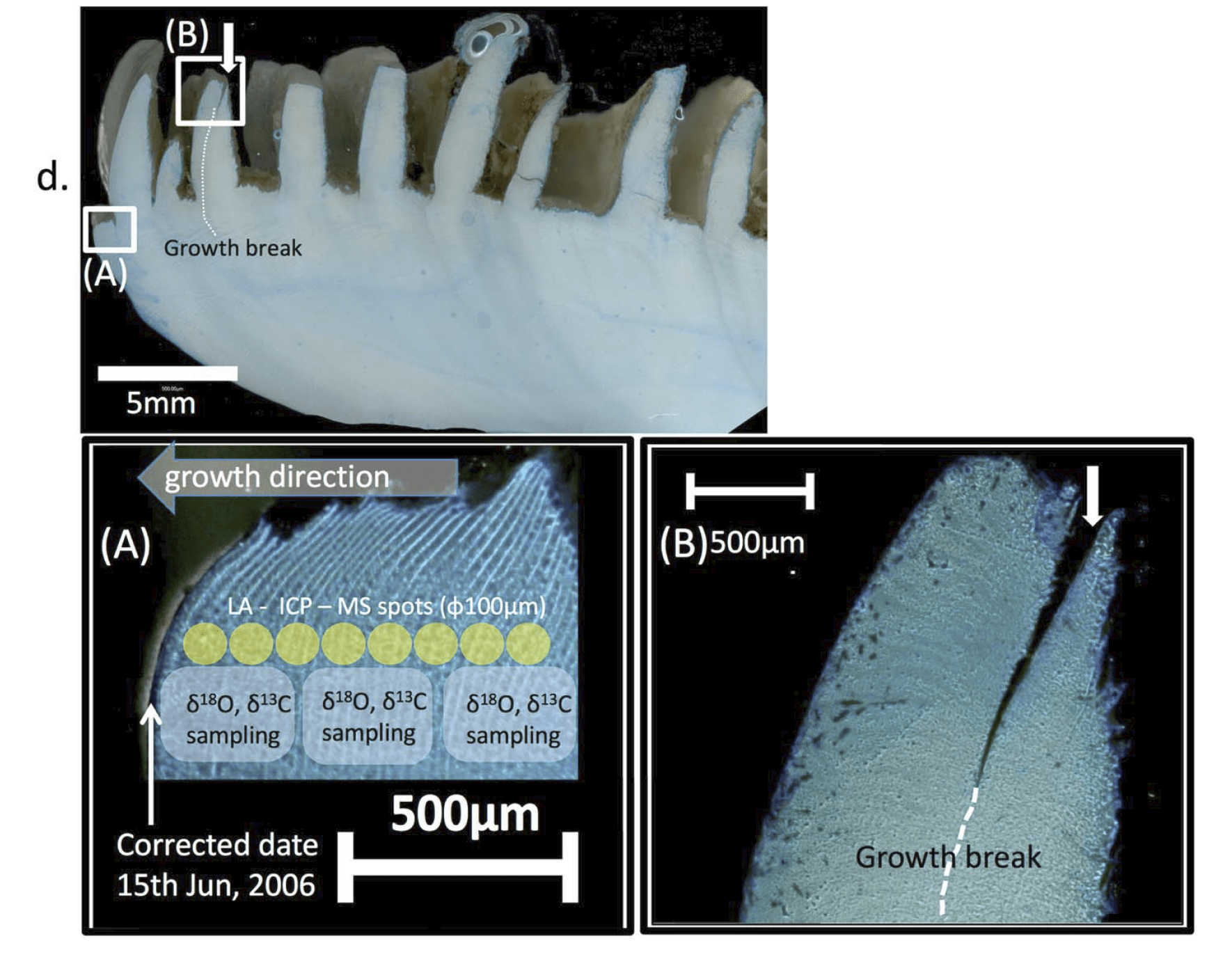
I am a sclerochronologist. That means I study the hard parts of animals, in this case the shells of bivalves. Like the rings of tree, bivalves make growth lines in their shells which can serve as a diary of their lives. Some of my past work has looked at using chemistry of the growth lines of giant clams to measure the temperatures they grow at, compare the growth of ancient and modern clams, and even look at how much the clams grow in a day! Today though, I’m talking about my most recent paper, which looks at how we can use the shells of giant clams as a food diary.
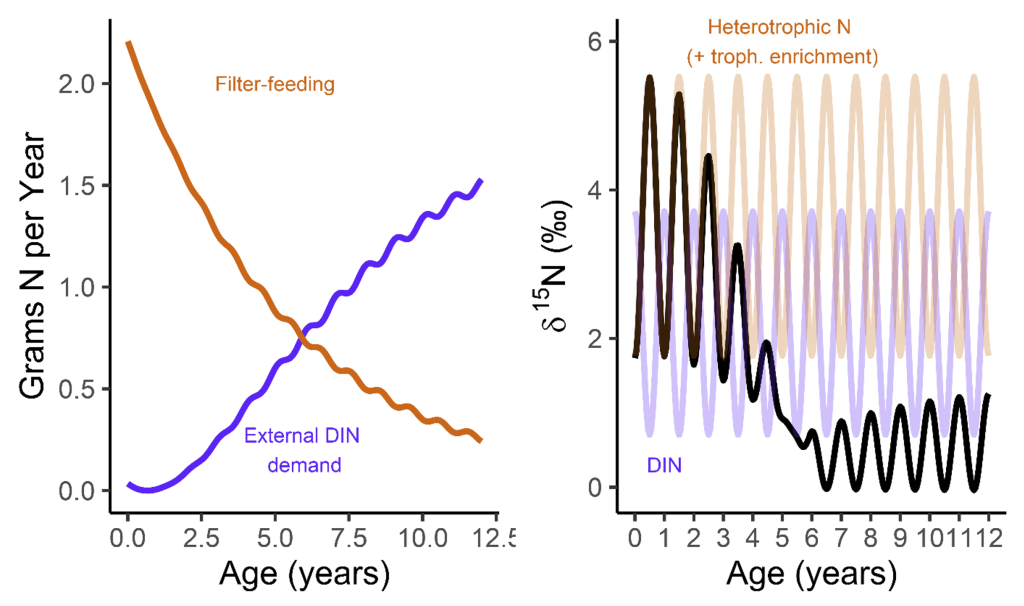
But when they’re babies, the symbiosis in giant clams is not yet fully developed. During this early period of their lives, giant clams actually get more of their nutrition from filter-feeding like a “normal” non-photosymbiotic clam, until they’ve had a chance to grow in surface area and become a living solar panel. Like all bivalves, the shells of giant clams are made of calcium carbonate, bound together by a protein scaffold we call the shell organic matrix. Proteins are made of amino acids, which contain nitrogen! If we can get the nitrogen out of the shell from the early part of the clam’s life, and compare it to the nitrogen at the end of the clam’s life, it might record the clam’s transition from filter feeding to its mature plant-like lifestyle! If our hypothesis holds, we should record a decrease through its life in the shell δ¹⁵N values.
A model I made of the clams’ nitrogen intake, with the left plot how they switch from filter feeding to getting most of their nitrogen from dissolved sources around 5-6 years of age. Because the nitrogen isotopes of those two sources are different, that manifests in the expected values from the clam’s body (the right plot)!
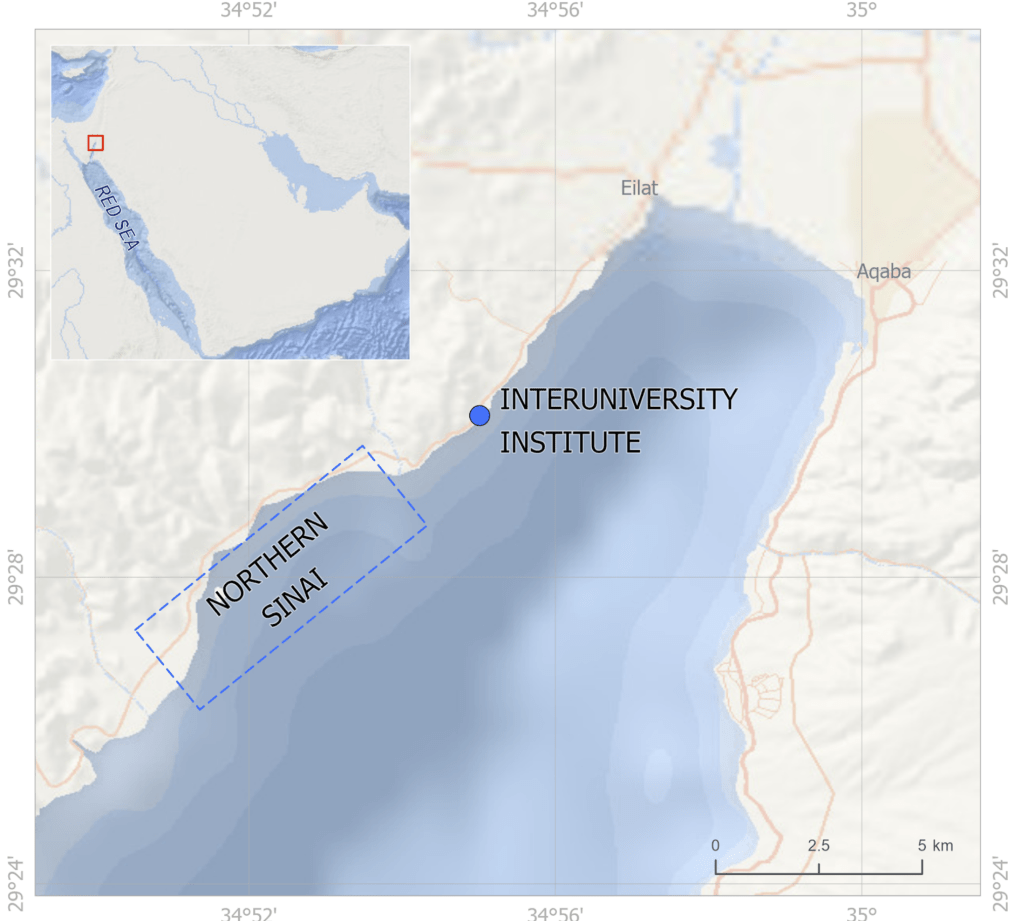
A map made by my talented partner, Dana Shultz!
So I gathered a team of talented collaborators and set out to test that hypothesis, using giant clam shells that I was able to get on loan from the Hebrew University of Jerusalem Museum. These shells had been confiscated from poachers at the Egypt-Israel border. While I would have rather known these clams were still alive in the waters of the Northern Red Sea, being able to use them for research to understand the biology of their species was the next best thing! I had originally planned on pursuing a postdoc undertaking this project with Rowan Martindale, a professor at UT Austin who has studied the nitrogen isotopes of photosymbiotic corals, but when I started up at Biosphere 2, we ended up continuing with the project anyway as a collaboration! We measured the nitrogen isotopes of the shell material in the lab of Christopher Junium, a professor at Syracuse University, who has developed an exquisitely sensitive method to measure the nitrogen from shell material by essentially burning the shell powder and then scrubbing out unwanted material to isolate the nitrogen, to measure the isotopes in a machine called a mass spectrometer. Katelyn Gray is a specialist in isotopes of biominerals and assisted with drilling out powder from the shells with a Dremel. Shibajyoti Das, now at NOAA, is a specialist in measuring the shell nitrogen isotopes of other bivalves and he was master at doing much of the mass spectrometer work, and assisting in interpretation. Adina Paytan is a professor at UC Santa Cruz. She first provided the funding and support for me to go to the Gulf of Aqaba and collect these shells as part of an NSF-funded student research expedition! She also provided environmental data which helped us to interpret what the clams were actually eating!
A figure showing the four shells we sampled from, with the sampling areas in each hinge area showing colored and matching with the corresponding isotope plot to the right (colored points). 3 of the 4 shells show declines in isotope values with age. Shaded ribbon behind the data shows the model output.
So what did our crack team of scientists find out? We found that three of the four tested giant clams did indeed measure a decline in nitrogen isotopes over the course of their lives. Their earliest growth lines in the hinge areas of their shells record elevated δ¹⁵N values similar to other filter-feeders from the region. But as they aged, their later growth lines show much lower δ¹⁵N values, more like photosymbiotic corals and plants from the region. So clams indeed recorded the transition in nutrition as they became solar-powered! This degree and directionality of change in nitrogen isotopes was much greater than has been observed in any other clams measured in this way, which made sense considering their unique physiology. The clams have another area of the shell, the outer shell layer, which is closer to the symbionts than the hinge area. In this outer shell area, we did not observe much of a consistent trend in nitrogen isotopes. It’s likely that the outer layer is highly influenced by the photosymbionts even at the earliest stages of life.
Growth lines in the hinge area of two of the shells lit from behind, with the drilled areas for this study visible as well. The outer shell layer is the opaque and was also sampled for this study.
There was one clam that differed from the others in showing low δ¹⁵N values through life in its hinge shell layer. To help explain these differences, I created an independent model of the clams’ internal chemistry based on their growth rate, which slows as they age, and also is faster in the summer. When the clams are young filter feeders, they get most of their nitrogen from plankton, debris and other material floating in the water column making up floating material we call Particulate Organic Matter (POM). Meanwhile, when they are in their photosynthetic life stage, they get most of their nitrogen from nitrate, which is essentially Miracle Gro for the symbionts. The model showed that the clams should record a flip from filter feeding to photosynthesis around 4-5 years of age, which was confirmed by three of the shells! But what about the one that didn’t show this trend? My colleague Adina had fortunately measured the isotopes of POM and nitrate in different seasons in the Gulf of Aqaba. We found that in summer, as expected, POM δ¹⁵N is lower than nitrate. In the winter, meanwhile, that relationship is flipped! So if a clam grew more in winter, it would not record the same transition as was seen by the other clams. We think the clam that was the exception to the rule might have been more of a winter grower.
The chaotic nutrient environment of the Northern Red Sea, showing how in different seasons, dissolved nitrate has higher or lower δ¹⁵N values than the Particulate Organic Matter that the clams filter-feed on.
But long story short, we were able to demonstrate for the first time that giant clams show nitrogen isotopic values in their shells in line with expectations from their diet. Other clams have been measured this way, but the fact that we were able to conduct these analyses at all is a testament to the sensitivity of the elemental analyzer in Chris’s lab. Giant clams have *very* low concentrations of organic matter in their shells, so the forward march of technology was a major factor enabling this study to be possible.
Why does it matter that we can measure the transition of the clams from filter-feeding to photosymbiotic in their shell records? Well, giant clams are not the only bivalves which have photosymbionts. There are other clams in the fossil record which have been proposed to have had symbioses with algae, but until now we’ve never had a definitive geochemical way to measure this in fossils. We hope that this approach can be applied to the organic material in fossil shells, which is often well preserved, to see if huge clams in the Cretaceous and Jurassic had a similar way of life to the modern giant clams! If we can demonstrate that was the case, we can see how such species responded to past intervals of climate change, which will help us understand how giant clams will fare in the warming, acidifying ocean of the present.
These results also help explain the lives of giant clams themselves. We hope this kind of data can be used to measure the symbiotic development of giant clams in different places, with different types of food and nitrogen available, where we’d have the potential to measure pollution. Interestingly, the time that the model shows the clams transitioning to photosynthetic maturity is right around the time that they reach reproductive maturity (5-10 years of age). We’d like to investigate whether the time of clam maturity is controlled by the development of their symbiosis, which itself might relate to nutrients in the clams’ environment. If clams can grow faster, then they can mature faster, and potentially reproduce sooner in life. Will giant clams be able to thrive in the presence of increased nitrate, which is a common pollutant in coral reef environments? Like all worthwhile research projects, we have dozens of new questions to pursue as a result of this work, so stay tuned for the next installment in this journey of clam knowledge!
The tree of life is often portrayed as a neatly branching structure, with each division point cleanly delineated and separated from its neighbors. The truth is that the various twigs of the tree of life often overlap and become tangled in a process we call symbiosis. I’ve talked about symbiosis before on this blog, which falls along a spectrum of wholesomeness. At one end we have mutualism, a partnership where both organisms benefit and achieve more than the sum of their parts. The other extreme is parasitism, when one organism benefits at the expense of the other. Between the two, there is a broad gray area including commensalism, when one organism’s presence doesn’t necessarily cost or benefit the other in any way. The tree of life is crowded and unpruned, and so sometimes the twigs might wrap around each other quietly and without much fuss. We live on a small planet, and have had to get used to living in uncomfortable intimacy with all sorts of creatures, such as the mites that are living on your eyelashes right now.
But things start to get really weird and tangled when the tree of life loops over on itself twice, or three times, or more. “Three-way” symbioses are surprisingly common, and the more you look for them, the more you realize that the tree of life is more of a knot than anything else.
A view of the Heteropsammia coral from the side
A recent paper from researchers in Bremen (Germany) and Saudi Arabia looked at such a three-way symbiosis between a coral, a worm and bivalves found off of Tanzania in East Africa. The relationship between solitary corals (Heteropsammia cochlea and Heterocyathus aequicostatus), a sipunculan worm (Aspidosiphon muelleri muelleri) and the clam Jousseaumiella, is a complex triangle of dependencies that had previously been noticed by other researchers, but never investigated at great depth. The worm lives with multiple tiny clams attached, all inside of a small solitary coral the size of a dime (1 cm long). Is the coral a willing host for this crowded boarding house, or has it been parasitized? Does the worm gain anything from the clams? The researchers sought to find out.
Part of the reason I enjoyed reading this study so much was that it had to take a narrative structure to describe the evolutionary ménage à trois of its focus. So much of modern science has moved away from anecdote to hard data, and while there is plenty of that to find in the study, it turns out that a lot of the study of symbiosis is storytelling. We need to know the setting and the characters.
In this case, the main characters are small solitary corals living in the tropical reefs of the Indo-Pacific. We denote them as solitary to distinguish them from their giant colony-forming compatriots that construct the coral reefs currently threatened by climate change and pollution. But like those giant reef-builders, these solitary corals get much of their food from sunlight through a mutualistic partnership with algae called Symbiodinium. The algae provide the host with sugars and other photosynthetic products, and the hosts give them nutrients and a safe cozy home in their tissue.
You might be thinking, “Wait! Dan just said this was a three-way partnership between a coral, a worm and some clams. So this is actually a four-way partnership between corals, worms, clams and algae?” You’d be exactly right. And I’m happy to say that the plot of this sordid story is about to thicken even further.
The side of the coral. See the little pores?
The Aspidosiphon worm is found in a spiral-shaped burrow inside of the skeleton of the coral. It is a pretty cozy home, with walls made of calcium carbonate by the coral, with breathing holes in the sides to allow the worm to breathe and release waste. The researchers wanted to know more about the structure of the burrow. Was it dug out by the worm using acid or an abrasive motion, like some clams use to dig into coral? So the researchers essentially gave the coral a CT scan to see its 3D internal structure. Inside they found growth features suggesting the coral grew around the worm, as if intentionally providing it a home.
Cross-sectional CT scans of the coral skeleton. In figure D, you can see the silhouette of the chambers of the snail shell where the worm made its first home!
Even more crazily, they found evidence that the worm had first settled inside an empty snail shell, like a hermit crab! The coral probably settled on a snail shell as a larva, and grew to engulf the whole snail shell, leaving growing space for the worm inside, with windows and all! So to review, this is now a five-way symbiosis between a dead snail, a worm that moved into its empty shell, the coral (powered by algae) that grew around it and encased the snail shell within its skeleton, and we haven’t even gotten to the clams. How many creatures are hiding stacked in this trench coat? Please bear with me as I explain!
An SEM image of Jousseaumiella. These are less than 1 mm long! Pinhead sized!
What are the clams doing in this picture? Jousseaumiella is part of a family of clams called Galeommatidae, which we previously mentioned on this blog in the context of some bivalves found growing in the gills of unfortunate sand crabs. Many members of the Galeommatidae family are parasitic or commensal with other marine organisms. In this case, Jousseaumiella are tiny flat-bodied clams less than 1 mm long, found attached to the body of the worm, squeezed inside the burrow in the coral’s skeleton. It feeds on the worm’s waste and potentially food particles coming through the pores in the sides of the burrow. Not the most dignified existence, but a more mobile home means more opportunities to eat a varied diet similar to that that the worm and coral are seeking out, and the clam also gets protection from predation tucked inside the coral. It is unclear if it benefits the worm directly to have clams attached to it.
A time lapse of the coral+worm moving from the paper’s supplement! It would be handy to navigate to greener pastures, if it became too muddy in a certain place!
It is, however, clear how living inside a coral would be a pretty good deal for the worm, which gets a stable, protective suit of carbonate armor to protect it from predators, and grows to fit it as it gets larger. They are normally found inside of rocks, shells and other hard inanimate objects, but having a living home is a cool upgrade. What is the coral getting out of the deal? The researchers note that the corals are often found in the crevices between other large reef-building corals, in areas of the reef that receive high supplies of nutrients and turbidity (dirt that blocks out light). These sorts of environments aren’t necessarily friendly places for a coral to be, since they reduce the light and therefore the food that the coral can receive from photosynthesis. These crevices also have a lot of variability in other conditions like temperature and water flow. But because the coral has hitched a ride on the back of a worm, it can actually move in the sediment to react to changing conditions and avoid being buried by piles of sediment floating by! The worm can also act as a sort of anchor preventing the worm from sinking in the sediment underneath, which would be a big hazard for the small, stubby coral on its own. The coral seems to go to great pains to make its partner comfortable, not growing its skeleton to cover the pore windows to the outside. The researchers note that as coral reefs worldwide are subject to increasing human-made pollution and climate change, it would be interesting to research whether this complex three-(five?) way symbiosis provides the various participants with an advantage compared to other corals.
So like any good story, this symbiosis features complex, growing characters, a dynamic setting, and still plenty of mystery demanding a sequel! To that end, there are lots of other great three-way symbioses to investigate. Snails which farm fungus that parasitizes plants. Bryozoans living on snail shells that have a hermit crab inside. Gobies serving as lookouts at the entrances of burrows built by shrimp, with a crab freeloader along for good measure. Algae and bacteria teaming up to attack mussels. The list keeps going! I could see this becoming quite a franchise!
When I mention to people that I study bivalves, I can sometimes sense from their facial expressions that they are secretly asking “why?” While clams are perfectly content to keep doing what they’re doing without being thanked, I think it’s important to enumerate all of the ways they make our world more livable and functional.
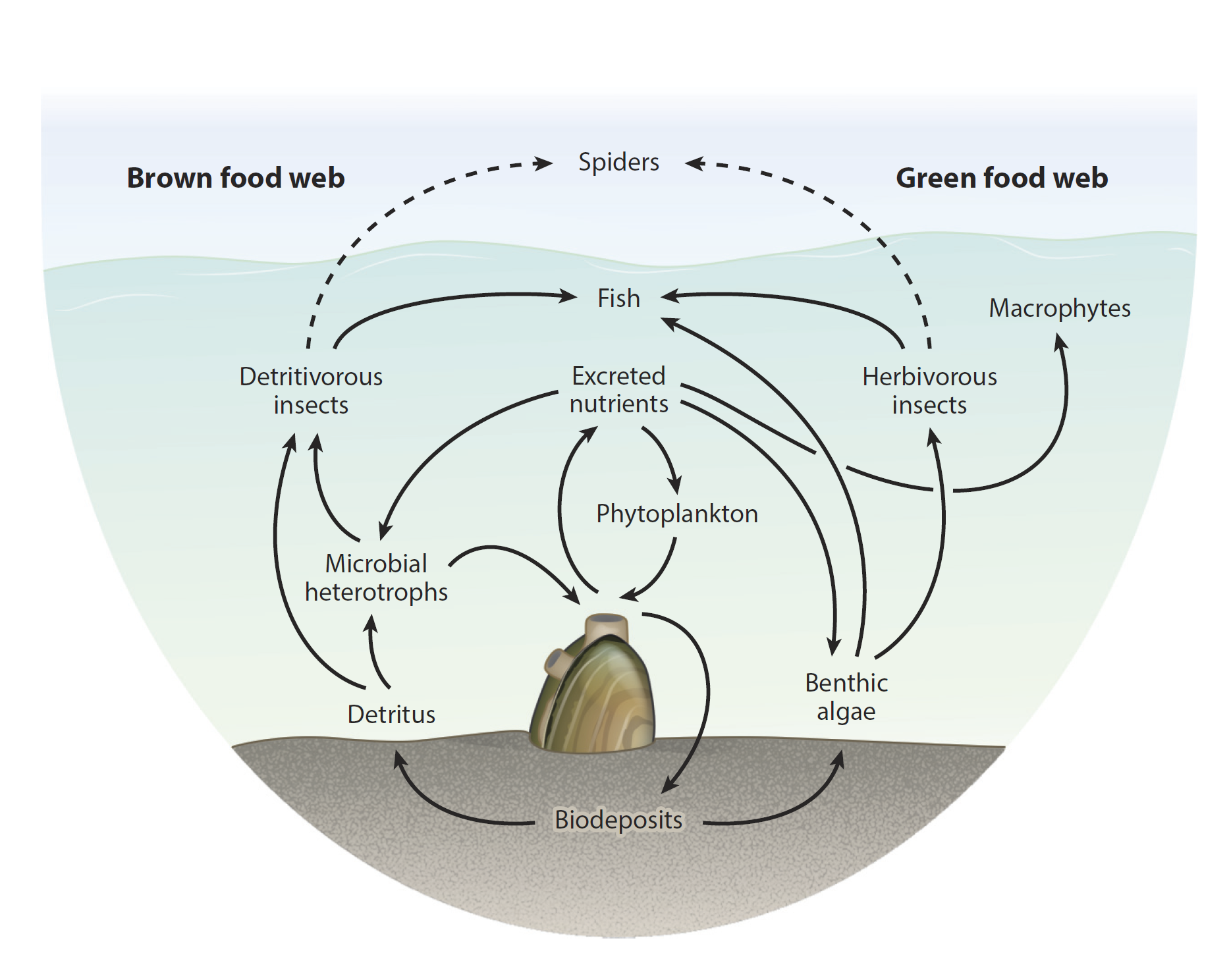
Various roles that freshwater mussels can play in their local food webs (Source: Vaughn and Hoellein, 2018)
Bivalves are ecosystem engineers. While they may seem rather stationary and not up to much at any particular time, they are actually always working to actively maintain their habitat. The majority of clams are filter-feeders, meaning that they use their gills to gather particles from the water column for food. Some of these particles are ingested as food and later pooped out. Some inedible particles are discarded immediately by the clam as “pseudofeces”. Both mechanisms serve as a bridge between the water column and the benthos (the sediment at the bottom). In this way, clams are engines that take carbon fixed by algae floating in the water and transfer that material to be stored in the sediment. Their bodies also act as nutrition to feed all sorts of animals higher on the food chain like sea stars, lobsters, seabirds, sea otters and humans that depend on bivalves as food. They are literally sucking up the primary productivity (algae) to be used by the rest of the food chain.
The filtration rate of oysters. Graphic from The Nature Conservancy
Different clam species vary in their precise filtration rate (how fast they can inhale and exhale water, filtering the particles within), but it is prodigious. Some freshwater mussels, for example, can pick-through 1-2 liters of water per hour for every gram of their own flesh. Since these individual bivalves can weigh over 100 g, they are capable of picking the food out of an immense quantity of water. In doing so, bivalves help improve the clarity of the water column, allowing more sunlight to reach deeper into the water body (the photic zone), providing more energy for additional photosynthesis to occur. While there are examples where invasive bivalves such as zebra or quagga mussels take this phenomenon too far, in well-functioning ecosystems, the filtration activity of clams helps improve the productivity of the community.
An oyster reef. Source: The Nature Conservancy
Bivalves help make sediment through their filtration of material from the water column, and they also engineer and manipulate the sediment directly. Some bivalves, like oysters, are able to make huge mounds of dirt that serve as habitat for all sorts of life, increasing the diversity of the community. They do so both by excreting sediment, and also by passively trapping it between the shells of neighboring oysters (“baffling”). By doing so, they reduce rates of coastal erosion and increase the biodiversity of wetlands. For this reason, New York and other communities plan to seed oyster reefs to help fight sea level rise and reduce the threat of storm surges like the one that occurred during Superstorm Sandy.
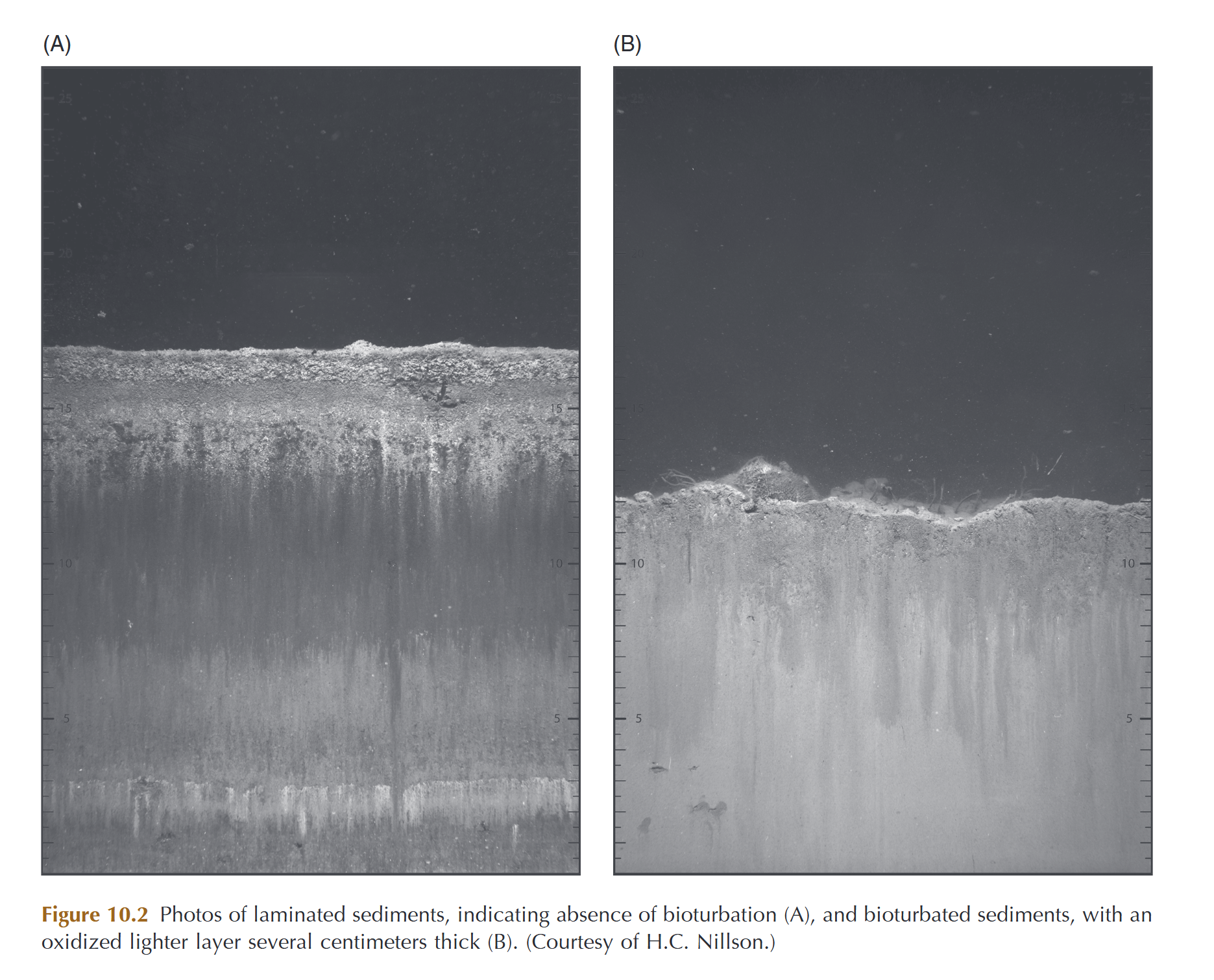
Comparison of sediments without bioturbation by digging animals, and with. Notice how the non-bioturbated sediment is layered and darkened due to activity by anaerobic bacteria, while the well-oxygenated, mixed sediment is light all the way through. From Norkko and Shumway, 2011
Other “infaunal” bivalves (burrowers) help to aerate the sediment through their tunneling, bringing oxygen deep under the surface of the dirt. This mixing of the sediment (also called bioturbation) ensures that nutrition from deep under the sediment surface is again made available for other organisms. Some bivalves can bore into coral reefs or solid rock, creating burrows which serve as habitat for other animals and can free up minerals for use by the surrounding ecosystem. Helpful shipworms assist in eating wood, assisting in returning nutrients stored in that tissue to the ecosystem as well.
Enormous grouping of giant clams in a lagoon in French Polynesia. From Gilbert et al., 2005
Bivalves of course are also famous for their shells, and this activity also provides habitat to sponges, snails, barnacles and many other encrusting organisms specially adapted to live on bivalve shells and found nowhere else. Giant clams are the most legendary “hypercalcifiers,” and in some regions like New Caledonia can rival reef-building corals in terms of biomass. In areas where soft-bottoms dominate, bivalves like hammer oysters, adapted to “rafting” on the quicksand-like surface of the soft sediment, can assist by providing a platform for other animals to take refuge. In the deep sea, bathymodiolid mussels and other chemosymbiotic bivalves can feed directly on the methane and sulfur emitted from hot vents or cold seeps with the help of symbiotic bacteria, creating dense reefs which provide food and habitat for all sorts of life. Even once the clams die, their shells can continue to serve as homes for other creatures.
Crabs feeding on Bathymodiolus in the deep sea (NOAA)
The shells of clams provide great scientific value in understanding our world. Much like tree rings serve as a record of environment thousands of years into the past, growth rings in clam shells serve as a diary of the animal’s life. These rings can be yearly, lunar, tidal or even daily in rhythm, with each ring serving as a page in the diary. The chemistry of those “pages” can be analyzed to figure out the temperature the clam experienced, what it ate, whether it suffered from pollution, and even the frequency of storms! The study of rings in the hard parts of animals is called sclerochronology, and it’s what first drew me to study bivalves. I was so fascinated by the idea that our beaches are covered with high-resolution records of the ocean environment, waiting to be cut open and read.
This giant clam shell recorded an interruption in the animal’s daily growth caused by a typhoon! From Komagoe et al., 2018
While they don’t owe us anything, clams provide a lot of value to humans as well, serving as a sustainable and productive source of food. Humans have been farming bivalves for thousands of years, as evidenced by “oyster gardens” and shell middens which can be found all over the world. Particularly in seasons when food is scarce on land, native peoples could survive by taking advantage of the wealth of the sea, and bivalves are one of the most plentiful and accessible marine food sources available. But they aren’t just the past of our food; they may be part of the future. Bivalves are one of the most sustainable sources of meat known, requiring very little additional food to farm and actively cleaning the environment in the process. Mussels grown out on a rope farm are an easy investment, growing quickly and with very little required energy expenditure. Someday, giant clams may provide the first carbon-neutral meat source, as they gain their food from symbiotic algae within their flesh. I have never eaten one, but I’ve heard they’re delicious.
A shell midden in Argentina. Photo from Mikel Zubimendi, Wikipedia
Mussels being farmed on ropes
Clams are heroes we didn’t know we needed and maybe don’t deserve. They ask for nothing from us, but provide vast services which we take for granted. So the next time you see an inconspicuous airhole in the sand, thank the clam that could be deep below for aerating the sediment. The shell of that long-dead mussel at your feet may have fed a sea star, and now is a home for barnacles and many other creatures. While that mussel was alive, it sucked in algae to improve water quality on our beaches. And the sand itself may contain countless fragments of even more ancient shells. Clams silently serve as an important cog in the vast machine that makes our oceans, rivers and lakes such amazing places to be. Thank you clams!
To us active, dynamic mammals, the humble clam can appear positively…inanimate. Their nervous system is decentralized relative to ours, lacking any sort of brain, and to the untrained eye, it can appear that their only discernible reaction to the outside world is opening or closing. Open = happy, closed = not happy; end of story, right? Some vegans even argue that the clams are so nonsentient that it is okay to eat them and think of them as having no more agency than a vegetable!
You might already have predicted I intend to tell you about just how animate and sentient clams can be. But let’s start out by describing the nuts and bolts of their nervous system. As with many invertebrates, their nervous system is distributed throughout their body as a system of ganglia. Ganglia are clumps of nerve cells which may have local specialization, and transmit messages within neurons using electrical potentials. At the connection between cells (called a synapse), neurotransmitters are used to pass signals to the next cell. Researchers have found that bivalves use “histamine‐, octopamine‐, gamma‐aminobutyric acid‐ (GABA)…like immunoreactivity” in their central and peripheral nervous systems, much like us vertebrates do, and other studies have even found that the response to serotonin and dopamine is localized in nervous tissue linked to different organ systems.
Nerve cells (bright green) highlighted in a larval oyster with fluorescent dye (from Yurchenko et al 2018)
These systems of chemical nerve transmission are truly ancient, likely dating back to the formation of complex animal body plans in the earliest Cambrian. Researchers have great interest in studying these nervous and hormonal signaling systems in mollusks because they can shed light on the relative flexibility and limitations of these systems throughout the animal tree of life. Characterizing these systems can also allow us to understand the mechanisms that bivalves and other animals use to react to environmental stimuli.
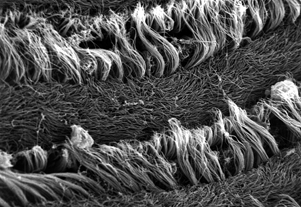
Electron microscope view of gill cilia, zoomed in 1000x (from Dan Hornbach)
Like humans, bivalves spend a lot of time and effort eating. Most bivalves eat by filtering food from passing water with tiny cilia on their gills. These cilia work to capture food particles and also act as a miniature rowing team moving water along the gill surface. The bivalve needs a way to control this ciliar activity, and researchers found they could directly control the speed at which oysters move their cilia by dosing them with serotonin and dopamine, which respectively increased and decreased activity.
Bivalves also work very hard to make babies. Most bivalves reproduce by releasing sperm and eggs to fertilize externally in the water column. To maximize their chances to find a mate, they typically save up their reproductive cells in gonads for multiple months and release them in a coordinated mass spawning event. It appears that this process is controlled by hormonal releases of dopamine and serotonin. Researchers have determined that serotonin concentrations vary through the year, with mussels in New England using it to regulate a seasonal cycle of feeding in summer, followed storing of that energy for winter. During the winter when food is less available, they use that stored energy to bulk up their gonads in time for reproductive release in spring months, when their larvae have plentiful access to food and oxygen, ensuring them the best chance of survival. In recent decades, aquaculturists have learned to use serotonin injections to induce spawning in cultured clams, to ensure they will have a harvest ready at a certain time of year.
So bivalves are very sensitive to the seasons. How about shorter term sources of excitement? You might have observed this yourself through the clam’s most iconic activity: opening and closing its shell. Clams close their shells with powerful adductor muscles which pull the two valves together. A springy ligament at the hinge pulls the shell open when the muscles relax. Just like us, the clam needs to use nerve cells to signal the muscle to do its thing. In addition, two different sets of ganglia act to control the foot that some bivalves can extend to dig into sand, with one ganglion acting to extend the foot and the other causing it to contract. While clams don’t have a centralized brain with specialized regions for different uses like we have, this represents a sort of specialization of neural systems with a similar result.
This iconic gif is often shared along with the claim it shows a clam “licking” salt. It is actually using its foot to search for a place to dig. The salt was not needed.
When a certain neuron is used repeatedly, it can form a cellular memory allowing the organism to acclamate (ugh sorry) and moderate its response to a particular stimulus over time. Giant clams, for example, close their shells when their simple eyes detect a shadow overhead. This behavior can protect them from predation. When I conducted some of my PhD research, sampling body fluid of aquarium and wild giant clams with a syringe, I noticed that captive clams didn’t close up in response to my shadow overhead, while wild clams required me to sneak up and wedge their shells open with a wooden block to do my work. I suspected that after exposure to frequent feedings and water changes by aquarists, the clam had “learned” that there was no reason to expend energy closing its shell. Meanwhile, in the process of proving that our sampling technique was not harmful to the animal, I discovered that clams which detected my shadow would quickly reopen within seconds when I hid from them, while those that were stuck by a syringe would stay closed for minutes before opening and beginning to feed again. Makes sense!
Other researchers noticed this phenomenon as well. One group found that giant clams repeatedly exposed to shadows of different sizes, shell tapping and even directly touching its soft tissue began to habituate (become accustomed) to the stress, opening more quickly and staying open longer each time the stimulus occurred. Even more interestingly, they did not transfer that habituation between stress types; for example, the clams that saw a shadow again and again would still react strongly to a different stress like tapping its shell. This suggests the animal can distinguish between different threats along a spectrum of seriousness, with touching of tissue (similar to a fish pecking at its flesh) being the most serious threat with the most dramatic response.
Another study determined that larger giant clams stayed closed longer than smaller ones in response to the same threat. They proposed this was related to the greater risk large clams face as they have more tissue area vulnerable to attack. While the clams might not have made a “conscious” decision in the way we do as thinking creatures, they were able to place their individual risk in context and vary their response. This ability to tailor a response to different risk levels is a sign of surprisingly complex neurology at work.
Close up of the eyes of a scallop. Each is a tiny crystalline parabolic mirror (photo by Matthew Krummins on Wikipedia)
Scallops show some of the most complex bivalve behaviors. This relates back to their unique adaptations, including simple eyes that can resolve shapes and the ability to swim away from danger. Scallops have been found to discern between predator types by sight alone, closing or swimming away in response to crustaceans but not sea stars. When swimming, they are capable of using this vision to navigate to places where they can hide, such as seagrass beds. They even use their vision to gather food: in one of my favorite experiments, researchers showed scallops a video of food particles, and they opened their shells and started filter feeding in response, kind of like me salivating when I watch the Food Channel! It would be very interesting to compare the behavior of scallops in marine protected areas to those that can be freely harvested. Do they vary their behavior in response?
I hope I’ve made clear that while clams are not exactly intellectual powerhouses, their behavior is much more complicated than simply sucking up water and opening or closing their shells. Like us, they inhabit a complex environment that requires a multitude of responses. Their nervous systems have evolved to allow them to survive and adopt nuanced behaviors which they can vary on the fly, and which us “higher” animals are only just beginning to comprehend.
The streamlined shells of Gaimardia trapesina. Source: New Zealand MolluscaBivalves are not known as champion migrators. While scallops can swim and many types of bivalves can burrow, most bivalves are primarily sessile (non-moving on the ocean bottom). So for many bivalves, the primary method they use to colonize new territories is to release planktotrophic (“plankton-eating”) larvae, which can be carried to new places by currents and feed on other plankton surrounding them. Many bivalves have broad distributions because of their ability to hitchhike on ocean currents when they are microscopic. They don’t even pack a lunch, instead eating whatever other plankton is around them. But once they settle to grow, they are typically fixed in place.
Not all bivalves have a planktotrophic larval stage, though. Larvae of lecithotrophic bivalve species (“yolk-eaters”) have yolk-filled eggs which provide them with a package of nutrition to help them along to adulthood. Others are brooders, meaning that rather than releasing eggs and sperm into the water column to fertilize externally, they instead internally develop the embryos of their young to release to the local area when they are more fully developed. This strategy has some benefits. Brooders invest more energy into the success of their offspring and therefore may exhibit a higher survival rate than other bivalves that release their young as plankton to be carried by the sea-winds. This is analogous to the benefits that K-strategist vertebrate animals like elephants have compared to r-strategist mice: each baby is more work, and more risky, but is more likely to survive to carry your genes to the next generation.
Brooding is particularly useful at high latitudes, where the supply of phytoplankton that is the staple food of most planktrophic bivalve larvae is seasonal and may limit their ability to survive in large numbers. But most of these brooding bivalves stay comparatively local compared to their planktonic brethren. Their gene flow is lower on average as a result, with greater diversity in genetic makeup between populations of different regions. And generally, their species ranges are more constricted as a result of their limited ability to distribute themselves.
A bunch of G. trapesina attached to kelp. Notice the hitchhiking clams have in turn had hitchhiking barnacles attach to them. Freeloaders on freeloaders! Source: Eleonora Puccinelli
But some brooding bivalves have developed a tool to have it all: they nurture their young and colonize new territories by sailing the seas using kelp rafts. The clam Gaimardia trapesina has evolved to attach itself to giant kelp using long, stringy, elastic byssal threads and a sticky foot which helps it hold on for dear life. The kelp floats with the help of gas-filled pneumatocysts, and grows in the surge zone where it often is ripped apart or dislodged by the waves to be carried away by the tides and currents. This means that if the clam can persist through that wave-tossed interval to make it into the current, it can be carried far away. Though they are brooders, they are distributed across a broad circumpolar swathe of the Southern Ocean through the help of their their rafting ability. They nurture their embryos on specialized filaments in their bodies and release them to coat the surfaces of their small floating kelp worlds. The Southern Ocean is continuously swirling around the pole due to the dominance of the Antarctic Circumpolar Current, which serves as a constant conveyor belt transporting G. trapesina across the southern seas. So while G. trapesina live packed in on small rafts, they can travel to faraway coastlines using this skill.
The broad circumpolar distribution of G. trapesina. Source: Sealifebase
The biology of G. trapesina was described in greater detail in a recent paper from a team of South African researchers led by Dr. Eleonora Puccinelli, who found that the clams have evolved to not bite the hands (kelp blades?) that feed them. Tests of the isotopic composition of the clams’ tissue shows that most of their diet is made up of detritus (loose suspended particles of organic matter) rather than kelp. If the clams ate the kelp, they would be destroying their rafts, but they are gifted with a continuous supply of new food floating by as they sail from coast to coast across the Antarctic and South American shores. But they can’t be picky when they’re floating in the open sea, and instead eat whatever decaying matter they encounter.
Falkland Islands stamp featuring G. trapesina. Source.
The clams are small, around 1 cm in size, to reduce drag and allow for greater populations to share the same limited space of kelp. Their long, thin byssal threads regrow quickly if they are torn, which is a useful skill when their home is constantly being torn by waves and scavengers. Unlike other bivalves, their shells are thin and fragile and they do not really “clam up” their shells when handled. They prioritize most of their energy into reproduction and staying stuck to their rafts, and surrender to the predators that may eat them. There are many species that rely on G. trapesina as a food source at sea, particularly traveling seabirds, which descend to pick them off of kelp floating far from land. In that way, these sailing clams serve as an important piece of the food chain in the southernmost seas of our planet, providing an energy source for birds during their migrations to and from the shores of the Southern continents.
A fan mussel among the seagrass it calls home (Arnaud Abadie on Flickr)
The fan mussels (Pinna nobilis) are a species of enormous mussel which live in seagrass beds of the Mediterranean Sea. They can grow to nearly 4 feet long (though most are 1-2 feet in size at maturity), and live with most of their bodies protruding straight up out of the sediment, anchored down into the sand with long rootlike byssal threads which grow out of their rear hinge.
These mussels grow up to 20 cm per year, almost entirely in the vertical direction. As they gain in mass, their bodies start to sink in the sand beneath them, so it is believed this extremely fast growth rate evolved in order to stay above the sediment. It also helps them to remain elevated above the seagrass around them, where they can access passing phytoplankton and organic particles in the current.
Close up view of the hairlike byssus. I definitely am feeling some beard envy here. (Wikipedia)
Because they are exposed to the current like a giant fan, they need a very strong anchor. So they create huge quantities of byssal threads which root them down in the sand. These byssal threads are known as as “sea silk” and communities around the Mediterranean have used the silk to sew clothing for thousands of years. The material is extremely fine but strong, and has historically been of immense value as a result. Sea silk or sea wool is mentioned in writings of the ancient Egyptians, Greeks and Romans.
Unfortunately, the fan mussels are considered critically endangered due to overharvesting, pollution, climate change and destruction of their native seagrass habitats. However, they are now protected and active conservation efforts are underway. When the cruise ship Costa Concordia ran aground off of Italy in 2012, a community of fan mussels were rescued from a seagrass bed next to the wreck and moved to another nearby site. I hope someday to study the fan mussels because I find them to be a truly charismatic bivalve with many interesting mysteries still waiting to be uncovered about their unique lifestyle.
Huge pen shell I saw at the Hebrew University Museum in Jerusalem. My lens cap is only 6 cm to give you a sense of scale! The shells are fragile and easily break.
Some shells of the carnivorous genus Cardiomya. Notice the protuberance off one side, making space for the overdeveloped siphon they use to capture prey (Machado et al. 2016)
You might think of clams as rather pacifistic creatures. Most of them are; the majority of bivalves are filter-feeding organisms that suck in seawater and eat the yummy stuff being carried by the currents. This mostly means phytoplankton, tiny single-celled photosynthetic plankton which make up most of the biomass in the world’s oceans. Most bivalves could be considered exclusively herbivorous, but as I’ve learned happens throughout evolutionary biology, there are exceptions to every rule. We already talked about parasitic bivalves that have evolved to hitch a ride on other hapless marine animals. But there is an even more sinister lineage of bivalves waiting in the sediment: yes, I’m talking about killer clams.
Carnivory in bivalves has evolved multiple times, but the majority of known carnivorous bivalves fall within an order called the Anomalodesmata. Within that order, two families of clams called the Poromyidae and Cuspariidae have a surprising number of species which are known to eat multicellular prey.
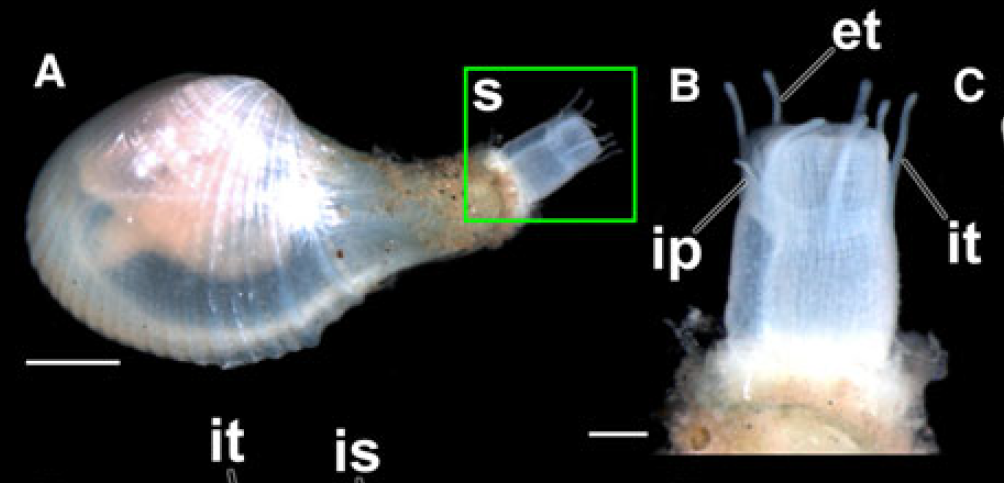
Now, you can rest easy because there are no clams that eat people. You’re safe from the Class Bivalvia, as far as we know. But if you were a small crustacean like a copepod, isopod or ostracod, you would be quite concerned about the possibility of being eaten by a poromyid clam in certain regions of the world. These clams lie in wait in the sediment like a sarlacc, with sensory tentacles feeling for passing prey and a large, overdeveloped siphon ready to suck up or engulf their helpless targets.
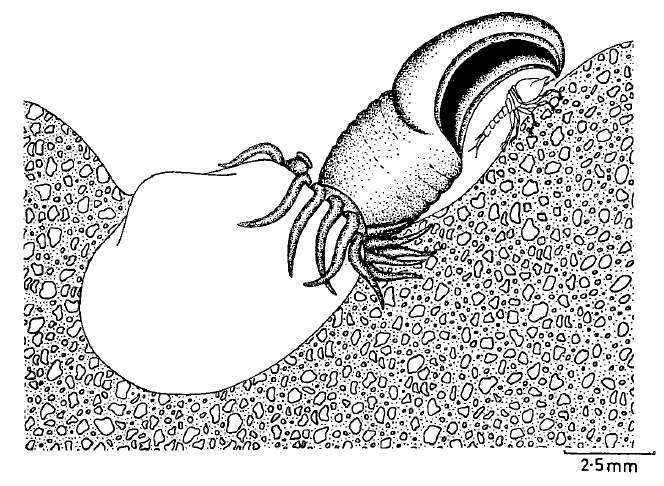
Until we catch the feeding behavior of poromyids on video, these whimsical artist’s depictions will have to do (Morton 1981).
Because they spend their lives under the sediment, these clams aren’t very well studied, and the first video of them alive was only taken in recent years. In addition, many of these killer clams live in deeper water, where their murderous lifestyle provides an advantage because food supplies can be much more sparse than in the sun-drenched shallow coastal zone. Much like the venus flytrap and carnivorous plants have arisen in response to the low nutrient supply of boggy swamp environments, the ability to eat alternative prey is valuable to the killer clams in all sorts of unconventional environments.
The siphon which these clams use to suck up their prey is a repurposed organ. In most other bivalves, the siphon is usually a snorkel-like organ which enables the clam to safely remain buried deep in the sediment and still breathe in oxgyen and food-rich water from open water above. But for the poromyids, the siphon is instead a weapon which can be used like a vaccum cleaner hose, or even be enlarged to engulf hapless prey. The poromyids have also evolved to have a much more complex, muscular stomach than any other bivalves. It takes a lot more energy to digest multicellular food, while most other bivalves simply just feed from the single-celled food they catch on their gills, expelling the other un-needed junk as “pseudofeces.”
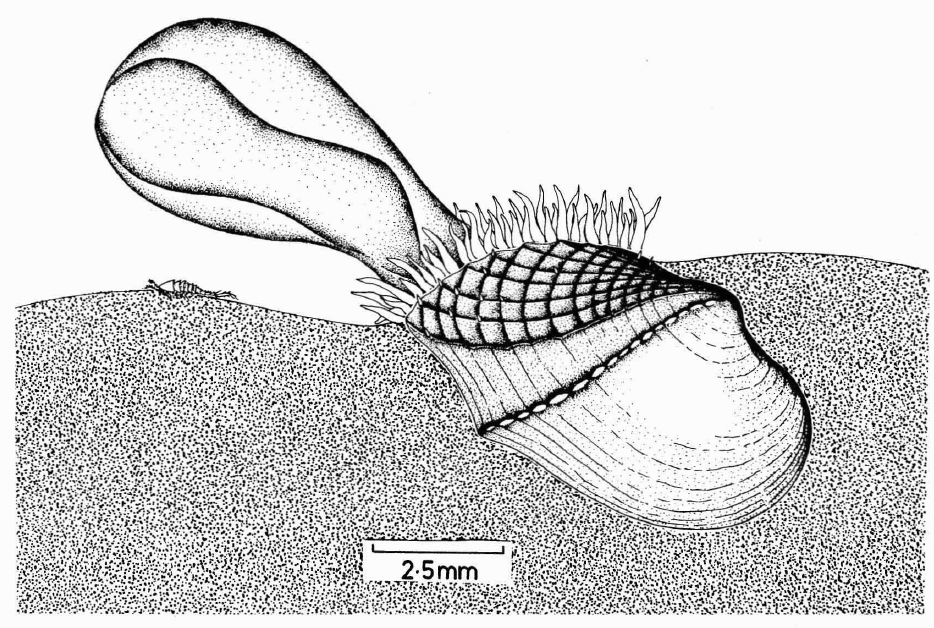
Dilemma, another strange carnivorous bivalve which eats marine isopods (pill bugs), found from deep waters off the the Florida Keys, Vanuatu and New Zealand (Leal 2008)
Hopefully soon we will have video of this predatory activity in action. But until then, you can imagine that somewhere on earth, tiny copepods foraging on the surface of the sediment pass by a strange field of squishy tentacles. Suddenly, out of nowhere a hellish giant vacuum hose appears in view and sucks them in like Jonah and the whale. Then it’s just darkness and stomach acid. What a way to go!
Lyonsiella going after a doomed copepod (Morton 1984).
Oyster. Reading that word, you probably formed an image in your mind of a rough-shelled creature with a shiny mother-of-pearl (nacreous) inside that someone pulled out of some silt in an estuary. And yes, that’s what most oysters look like. Some oysters are of additional economic value through their creation of pearls. These pearl oysters have long, straight hinge lines and live in the tropics in and around coral reefs.
A pearl oyster. See the straight hinge? Source: Pearl Paradise on Flickr
The hammer oysters are another sort of oyster, not of the Ostreidae family that includes most of the bivalves we think of as oysters, but still closely related and in its own family, the Malleidae. Malleus is the latin word for hammer, and the most distinctive genus of hammer oysters indeed look just like a hammer sitting on the seafloor.
In a typical life position in a seagrass bed. Notice all the algae, anemones and other encrusting creatures freeloading off the hammer oyster’s hard work. Source: Ria Tan on EOL
What the…that thing’s alive? How does that even work? This is an oyster? That’s how I imagine the first scientist to discover the hammer oyster reacting. Because they are weird and rather incomprehensible-looking. But when you know the way they live, it makes more sense.
There is a small area of nacre (mother of pearl) in the area near the rear of the interior. Source: Archerd Shell Collection
The hammerhead part of the oyster is just a super elongated hinge. The creature has a long, straight hinge like other oysters, but it has evolved to instead have a relatively narrow set of valves attached to that ridiculously overbuilt hinge. Like other oysters, they secrete byssal threads from their backside to attach themselves to the bottom. The narrow valves commonly poke up out of sandy bottoms in tropical waters nearby coral reefs. They do particularly well in seagrass beds, and often live in large colonies similar to other oysters.
Shell collectors seek out hammer oyster shells which have other bivalves attached. Here is a thorny oyster living on top of Malleus. Two for one! Source
The absurd hinge helps these creatures to stay anchored into the sediment, but also serves as “wings” that help it avoid sinking into the sediment over time. One thing us humans don’t realize sitting on sand is that it actually acts like a liquid. Over time, if we sat on wet sand, we would likely begin to sink in unless we spread out our arms and legs to increase our surface area. In the ocean, all sand is quicksand. Different organisms have different strategies to avoid being engulfed by the sediment they live on, and the hammer oyster has had good success with its strategy. It doesn’t care that you think it looks weird. It just sits there, filtering water for passing food particles and plankton. It’s very good at it, has been perfecting the strategy for over 250 million years, and doesn’t need your smartass remarks, thank you very much.
Another shot of a happy hammer oyster doing what it does best, in a seagrass bed near Singapore. Source: Wild Singapore on iNaturalist
The heart cockle (Corculum cardissa) is so named because of its heart shaped shell. It is native to warm equatorial waters of the Indo-Pacific. While many bivalves sit with the their ventral valve facing down, the heart cockle sits on its side, with one side of both valves facing downward. The valves have adapted to resemble wings and are flat on the bottom, providing surface area that allows the bivalve to “raft” on the surface of soft sandy sediment and not sink. They may also sit embedded in little heart-shaped holes on the tops of corals.
Two heart cockles embedded in the top of a Porites coral. Source: Reefbuilders
A particularly green heart cockle from Singapore. Source: orientexpress on iNaturalist
Heart cockles are a member of a small club of bivalves which partner with symbiotic algae for nutrition created by photosynthesis. Most of the modern photosymbiotic bivalves are in the family Cardiidae, the cockles. The giant clams (Tridacninae) are also in this family and have a similar partnership with the same genus of Symbiodinium algae. This algae is also found in many species of coral.
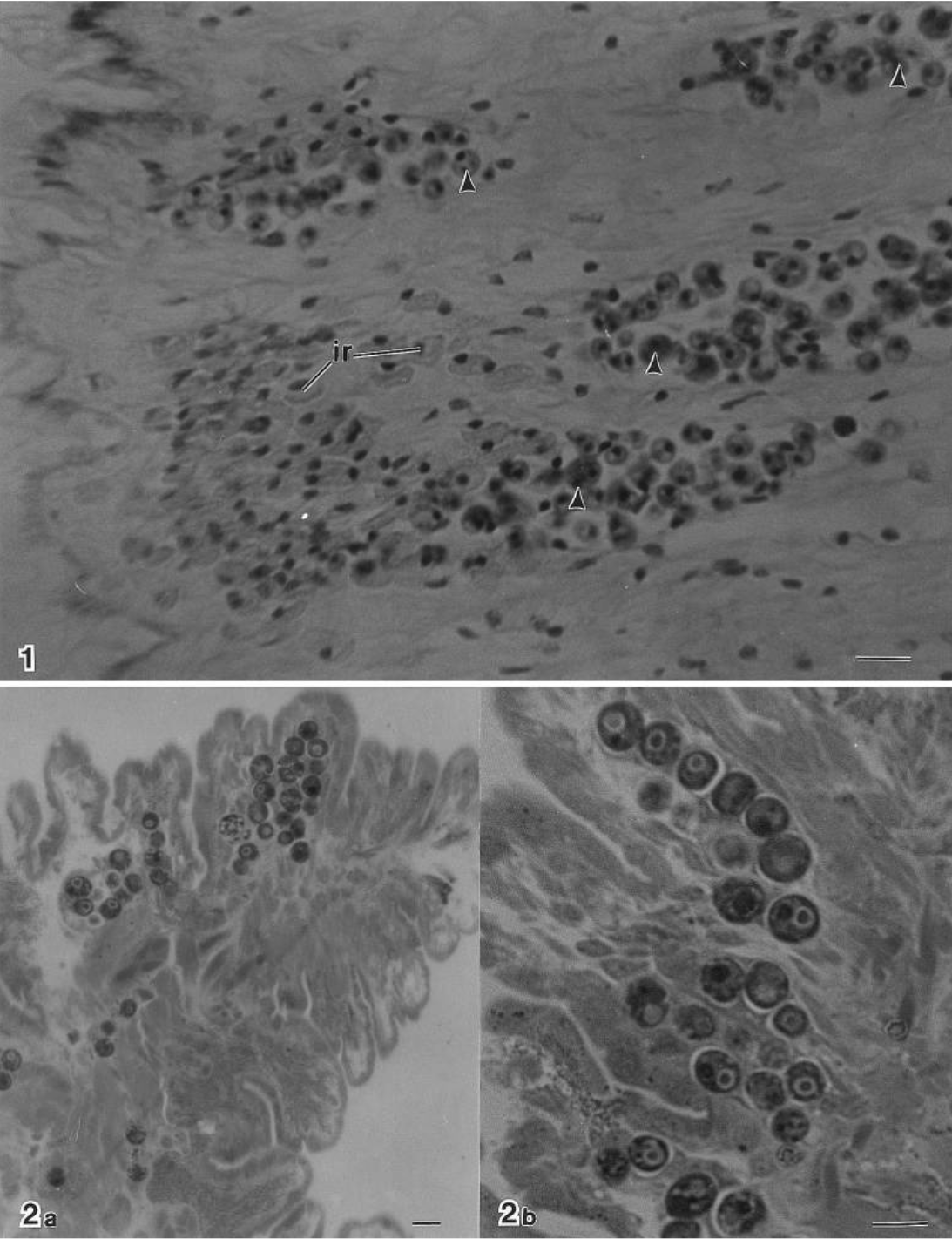
The dark circles in these microscope images are Symbiodinium. The top is a view of giant clam body tissue. The cells are present throughout the tissue in giant clams. The bottom shows heart cockle “tubules” which contain their symbiotic algae. The algae are restricted to narrow tubes that run through the tissue of the cockle. Source: Farmer et al. 2001
So when you find a live heart cockle, it is often green in color, because of the presence of this algae near the surface of its tissue. Its shell has adapted to be “windowed” (semi-transparent) to allow in light for the algae to harness to make sugars. The algae are housed in networks of tubes within the soft tissue of the cockle. They trade sugars with their host in exchange for nitrogen and carbon from the clam.
As I’ve mentioned before regarding the giant clams, this is a very productive partnership and has evolved separately several times in the history of bivalves. However, we don’t know why almost all examples of modern bivalve photosymbiosis occur in the cockles. Why aren’t the heart cockles giant like the giant clams? What features are necessary to allow this symbiosis to develop? These are the kind of questions I hope to help answer in my next few years of work.











































{kind=link}
{kind=link}
{kind=link}
{kind=link}
#/media/File:Naturalis_Biodiversity_Center_-_ZMA.MOLL.1264_-_Malleus_malleus_(Linnaeus,_1758)_-_Malleidae_-_Mollusc_shell.jpeg){kind=link}
{kind=link}