Screencap of Onion where some sort of scientist is announcing something in front of a Powerpoint slide of a clam
Last week, The Onion, a very serious journalistic publication, published a piece “Biologists Announce There Absolutely Nothing We Can Learn From Clams“. As a print subscriber I want to say I played a small part in this article, which I’ve actually hung on my office wall. But I want to take it a step further and write a line-by-line concurrence with everything they wrote!
WOODS HOLE, MA—Saying they saw no conceivable reason to bother with the bivalve mollusks, biologists at the Woods Hole Oceanographic Institution announced Thursday that there was absolutely nothing to be learned from clams.
Wow, I do know a researcher who studies clams at Woods Hole and actually love her work! Nina Whitney is now a prof at Western Washington University but until recently was a postdoc at WHOI studying how shells can serve as records of climate! I wonder who The Onion interviewed.
“Our studies have found that while some of their shells look pretty cool, clams really don’t have anything to teach us,” said the organization’s chief scientist, Francis Dawkins, clarifying that it wasn’t simply the case that researchers had already learned everything they could from clams, but rather that there had never been anything to learn from them and never would be.
Oh I don’t know a Francis Dawkins, but I’m sure they know their clams! It is true that their shells can look pretty cool. Bivalves include everything from Hysteroconcha dione, with its beautiful color and spines, to Tridacna gigas, which grows to 4.5 feet and weighs hundreds of pounds! And like an Onion, shells have growth layers, sometimes a new one every day, which someone could use to try to figure out how clams record what they eat and how the environment changes. But why would anyone do that?
Close-up view of a Hysteroconcha bivalve shell, showcasing its intricate ridges and coloration, and rows of long spines near the margin. Source
For me, I guess I haven’t learned anything from clams. I think I already knew in my heart that clams can live for >500 years. I already knew that mussels can filter several liters of water per hour, meaning that a colony of them can filter thousands of liters an hour. All this stuff is obvious, actually. Common sense.
“We certainly can’t teach them anything. It’s not like you can train them to run through a maze the way you would with mice. We’ve tried, and they pretty much just lie there.
It is ludicrous that clams could be taught anything or have anything approaching memory or thinking. It is only coincidence that scallops appear to clap their valves to swim, using their hundreds of eyes to navigate to a new location away from predators or toward food. It’s coincidence that they increase their feeding activity when shown a video of food particles. Some researchers have even claimed that giant clams can tell the difference between different shapes of objects! It is so dumb!
From what I’ve observed, they have a lot more in common with rocks than they do with us. They’re technically alive, I guess, if you want to call that living.
Also literally true! Their shell is a biomineral, in essence a living rock, made of calcium carbonate. They are alive in the sense they have a heart that beats, pushing hemolymph around their body. Their heart rate can increase or decrease with different stressors. Remember though, we always knew this. We didn’t learn it through something like science.
They open and close sometimes, but, I mean, so does a wallet. If you’ve used a wallet, you know more or less all there is to know about clams. Pretty boring.”
I myself have wasted time studying this. I attached sensors to giant clams to monitor their feeding activity. If I had learned anything, it might have been that they change their behavior between day and night, basking in the sun to help their photosynthetic algae in the day, and partially closing at night, with those behaviors changing based on how much chlorophyll is in the water. But remember! I didn’t learn it.
The finding follows a study conducted by marine biologists last summer that concluded clams don’t have much flavor, either, tasting pretty much the same as everything else on a fried seafood platter.
I can’t see how anyone would like to eat a bivalve. Especially not a fresh-caught scallop sauteed in butter or a plate of fried clams in New England. Never try that. Leave it to me!
Source: CDWR on Facebook. I thought about titling this post “Violence of the Clams” until I realized American Dad beat me to it, releasing an episode with that title in 2024
I legitimately admire clams. I whole-gilledly believe that they do a lot of good for the world; way more than we do! But there’s no doubt that some types of clams are up to no good, thanks to our help. One of those species is Limnoperna fortunei, the golden mussel. In late 2024, this species was observed for the first time on the North American continent, found attached to various human infrastructure in the Sacramento Delta of California. Since then, it has made its way down the California aqueduct all the way to the Southern tip of the Central Valley. Golden mussels are a notorious invasive species, and California officials immediately recognized the potential for disaster here, leading to dramatic policies of containment throughout the state that have tremendously impacted the lives of people trying to enjoy life on the water.
Map from CDFW showing the locations golden mussels have been observed as of July 2025
Since we are in uncharted waters with these mussels, there are a lot of questions about these innocuous-looking but trouble-making clams. In this blog, I will try to answer some of the most frequent questions I’ve seen over the last few weeks. I will caveat this by saying that I currently have no active research on this species, but I am a card-carrying clam scientist, and have a lot of interest in its biology and the significance its presence it will have for our state. So let’s get into it!
What are golden mussels? Where are they originally from? How did they get here?
A clump of golden mussels observed in Brazil by iNaturalist user danialdias
Golden mussels are small mussels, only reaching a bit over an inch in length, native to the Pearl River basin in China (the area around Hong Kong and Macau), but have been spread around the world over recent decades with the help of humans, hitching a ride between continents in the ballast water of our ships. Once settled in a new place, they easily move between lakes attached to boats being driven around, since they can live up to ten days out of water (talk about holding their breath!). The mussels first spread throughout Southeast Asia, then to Japan, then South America, and now for the first time, to the North American continent. While they are true mussels, in the same family (Mytilidae) as the more famous saltwater mussels you might have seen in tide pools, they can’t tolerate fully marine conditions.
Why are they a problem?
A now-infamous photo of the mussels coating the inside of a pipe at the Governor Jose Richa Power Plant in Brazil
They love to attach to heat exhangers, which can cause dams and pump stations to break down. Both are crucial to the California Aqueduct. Photo by Gustavo Darrigan
They can attach to native mollusks, smothering them. Photo by Gustavo Darrigan
Golden mussels are prolific breeders and make a living by anchoring themselves to any available hard surface using byssal threads. This is relatively uncommon among freshwater bivalves, most of which live on the bottom and don’t attach to surfaces. Golden mussels reproduce by releasing thousands of tiny larvae which spread through the area on river currents. In areas where they attach (such as dams, aqueducts, boats and other infrastructure), they form dense colonies that gum up the works, clogging pipes and and coating surfaces with thousands of their sharp little shells. They can even attach to the roots of native plants and shells of other molluscs and smother them! This causes hundreds of millions of dollars in damages and continuing expense in reservoirs and irrigation systems where they’ve taken hold, like in Japan and South America. If the mussels were to unexpectedly clog the outlet of a Californian reservoir like Lake Berryessa or Folsom Lake, it could be disastrous for people who depend on that water.
Figures from a paper about golden mussels invading Brazil, showing them coating an aquaculture cage, on a buoy, on a power plant hatch, in the entry to a dam turbine, and clogging a cooling pipe
Quagga and zebra mussels, originally from Central Asia, are invaders in the Colorado River, the Great Lakes, and reservoirs in Southern California. They have been limited from spreading into most reservoirs in Northern California by the low calcium content of lakes here (a function of our local rocks and geology). But golden mussels have lower calcium requirements than zebras/quaggas, so it is likely that they can reproduce in reservoirs up here. They are also surprisingly resistant to low temperatures, meaning that they could potentially take hold in high-altitude lakes like Lake Tahoe, which could be a disaster for efforts to keep Tahoe blue.
Why are they so successful?
A growing golden mussel colony, with an adult surrounded by younger babies. They only live about 3 years, but what a life they’ll live! Photo by Alexander Karatayev via Great Lakes Echo.
Being so prolific in their numbers allows the mussels to transform the chemistry and biology of the waters where they live. Like most bivalves, golden mussels make their living by using their gills to filter particles out of the water column, drawing them down to their mouth to eat. While individual golden mussels are pretty average in their filtering ability, together they work to much more effectively clear the water than other species, thereby depriving those native species of the plankton food they need, and potentially even directly eating the plankton larvae of other animals around them!
Figure showing how densities and size of colonies of toxic cyanobacteria Microcystis increase in the presence of golden mussels. Yum! Source
The Sacramento Delta has plenty of plankton floating around, so it’s not surprising they’ve decided this is a nice place to live. But while the water-cleaning ability of clams is a useful service they provide, there can definitely be too much of a good thing. The mussels are “ecosystem engineers”, meaning that they make the environment they want to live in. The problem is that what is good living for the mussels is not necessarily the habitat of a thriving Delta. Where they take hold, they exclude native species and generally decrease water quality by trapping dirt and boosting the populations of cyanobacteria. The Sacramento Delta already struggles with toxic cyanobacteria, and don’t need to have the problem be worse! Lower water quality means fewer fish, which is bad for people and the ecosystem.
Why have they shown up now?
A handful of Corbicula fluminea (Asian clams), a different species of invasive clams in the CA Delta. Photo source
This is actually not the CA Bay/Delta’s first rodeo with foreign clams. Invasions of Asian clams (Corbiculafluminea) and overbite clams (Potamocorbulaamurensis) in the 1980s transformed the Bay, with trillions of clams spreading out all the way south towards San Jose and eastward into the Delta after being introduced in Grizzly Bay in the mid-1980s. These clams had enormous impacts on the ecosystem, excluding other bottom-dwelling animals and eating most of the plankton food that other animals rely on. They are thought to have played a major role in the decline of some native fishes like Delta and longfin smelt.
Golden mussels have been making their way around the world over the decades. It is hard for their larvae to survive a couple weeks in the belly of a ship, be released, and successfully take hold, but with enough ships coming to California, it was only a matter of time before all of the stars aligned and a population took hold. We don’t know if the appearance of golden mussels will push out Asian clams, or if they’ll coexist. Asian clams live on the bottom rather than attaching to stuff, but golden mussels may still compete with them for food.
Are there other ways they spread?
Previous studies investigating their spread in South America and Japan determined that virtually all of their spread happens attached to the hulls of ships, in ballast water, or anywhere their larvae can travel downstream. There are rare cases where they are believed to travel upstream in the guts of fish that eat them, being pooped out alive. But those are unusual cases. That also won’t help them spread past dams without a fish ladder. The planktonic larvae have very little ability to swim against the current, so they won’t be able to swim upstream through dam turbines.
A map and timeline of their spread through Japan. Source
A map of their appearances and spread through South America. Source
Unlike pea clams, which are famous for attaching to birds by clamping their shells on their feet or feathers and traveling long distances to reach new places, it is not believed that golden mussels can create their thread attachment fast enough to hitch a ride on birds (which is a process that takes hours). So fortunately, I can assure our avian friends that we won’t need to inspect them before they use our reservoirs. At the end of the day, human vessels are the main way these mussels are getting around to far-flung places. In Japan, it took around 15 years to spread river to river through the country, while in South America, it covered most of a large area from Buenos Aires to Southern Brazil in that same period of time, which was proposed to be largely due to greater boat traffic in South American rivers.
Different life stages of larval golden mussels. They’re cute when they’re babies! The bottom right is the “plantigrade” stage when they attach to a boat, at 0.75 mm size. Imagine scanning a boat looking for one of those!
Are they good eatin’?
They don’t look exactly appetizing to me. Notice the visible byssal threads! Source: Folsom Lake Recreation Area
These mussels measure only a little over an inch at best, with not much meat on them. Unlike Asian clams (Corbicula), which are eaten in some Asian cultures, I can’t find mention of anyone eating golden mussels. There have been attempts using them as a fertilizer calcium supplement, but that needs more research. Additionally, it’s known that the other invasive clams of the Bay/Delta are concentrators of toxins, including selenium from farm runoff, heavy metals, and also toxins from harmful algae. In places where golden mussels colonize, toxic cyanobacteria can proliferate, so they actually make themselves a bit more toxic than other clams in the same place would be!
I don’t think these will be taking over the tapas restaurants any time soon! Source
What can we do about them?
We just don’t know how L. fortunei will fare long term in the California Delta and lakes. The previous clam invasions have waxed and waned through time. It’s uncertain whether these mussels will fizzle out, as sometimes happens for invasive species, or if they’re here for the long haul. The speed of their spread throughout the state personally leads me to suspect they’re here for good. And in the meantime, the invasion has caused huge issues for anglers, boaters and dam operators throughout California this summer, who have had to institute boat inspections at every reservoir in the state. Boats have to be painstakingly checked for mussels stuck to surfaces on the hulls.
Eventually, it is possible that mussels will find their way through, despite these precautions. Some could be missed in the crevices of boats entering various reservoirs. But hopefully that will buy time for dam operators to put forth the needed upgrades and develop procedures to keep them from fouling dams and aqueducts. At that point, the objective becomes mitigation rather than prevention. It won’t be cheap, usually involving manual scraping of mussels off of surfaces, application of hot water, pesticides, and use of surfaces that discourage mussel growth.
Map from a 2015 book chapter showing their distribution at that time on top, and predicted places they could invade on the bottom panel. Just as the prophecy foretold! Source
Long-term, our invasive species management needs to be more proactive rather than reactive. California was previously recognized to be in the range of territory where golden mussels could appear (see figure above). We can’t allow future invasions to catch us by surprise. To that end, there are laws on the books in California requiring inspection of 25% of incoming ships. So far, we are only inspecting a small fraction of that number. Additionally, ships were previously required to release ballast water far offshore in the ocean, where freshwater species wouldn’t be able to get a foothold. That policy was also not adequately enforced, and requirements to sterilize ballast water with chemical treatments were ruled too expensive. The state government very recently strengthened the standards, but gave ships until 2030 to comply with a weakened version of the rules, and pushed off compliance with the final strongest version until 2040!
People frustrated about such invasive species in California should insist to their policymakers that we can and must do better. There are manymoreinvasiveclam species waiting for their chance at a ride over here to make a living in our waters. It’s not too late to stop the assembly line of species coming to displace the native creatures we all love and value!
I often get asked what pearls are and why bivalves make them. Pearls are biogenic gemstones. This means they are valuable rocks made not by inorganic crystallization within the earth, like most gemstones, but instead are produced by life! Interestingly, they are living rocks, composed of true minerals. When I talk about minerals, I mean a solid substance with a known chemical composition and crystal structure.
Pearls are specifically made mostly of a mineral called aragonite. Aragonite is a mineral made of atoms of calcium bound to an ion called carbonate. There are other minerals made from calcium carbonate, like calcite and vaterite. I’ll save those for another blog, and while those are present in small amounts in some pearls, the vast majority of the material is aragonite. The clam uses calcium carbonate to make pearls because it’s a conveniently available material: it’s also what they build their shells from!
Aragonite has a very specific geometric crystal structure at the molecular level, but zooming out slightly, it can be found in a tremendous variety of microfabrics. Like the fabric of our clothes, the shell is essentially “woven” by the bivalve with a certain texture at the cellular level. There are hundreds of types of fabrics, ranging from structures looking kind of like brickwork, to plywood, to actual long fibers of carbonate. But the most valuable pearls are made of a form of aragonite called nacre, which is also called “mother of pearl” for this reason.
The platy microstructure of nacre. See how the tablets are organized into interlocking columns! Source: Wikipedia
Nacre is a very special biomineral for many reasons. To humans, it’s precious because of its beautiful, complex iridescence and luster, which has attracted our eyes for thousands of years. But most clams aren’t making the material for its luster- they value its microfabric. Nacre is made of billions of tiny flattened tablets of aragonite, arranged in tall interlocking stacks. Each aragonite tablet also has little bridges joining it to the neighboring tablet, meaning they don’t easily slide out of place. The plated structure also aids the shell in staying together. Even when fractured, the shell can stay together as plates slide to lock into another shape!
Reviewing the various strengthening aspects of nacre, including the bridges that lock tablets together, the rough surfaces of the tablets that grip against each other, the organics that glue together tablets like mortar, and the tablets sliding into new locking orientations even if they break apart! From Zhao et al., 2018
Between the bricks of aragonite are a kind of mortar or scaffolding of protein, binding them all together. This protein scaffolding is extremely important to the overall material. Like the steel rebar in concrete, it strengthens the material, making it less brittle and therefore able to resist forces that might crush the clam’s shell, while still allowing for the material to be very thin. For us humans, those alternating layers of carbonate and protein act like thousands of layers of prisms, refracting the light into thousands of colors depending on the angle it is looked at, meaning that any light becomes a miniature rainbow when it passes through the structure of the nacre.
A snuff box made from a nacreous bivalve shell, at the Vienna Natural History Museum
This structure makes nacre a “premium” material for clams to build their shells from. It costs much more energy for a clam to make such an orderly microstructure and fill it with so much protein, up to 5% by weight, which is around 5-50 times how much organic material is found in more common forms of aragonite. Nacre is also more vulnerable to dissolving in the water surrounding the clam. For this reason, clams usually will only use nacre in the internal shell layer, isolated from the surrounding waters, using cheaper materials on the outside of the shell, or at least a protective sheath of protein on top (called periostracum).
The nacre is present in mussels, oysters and other bivalves vulnerable to crushing pressures of waves as well as crushing predators like fish and birds. Nacre is like the bulletproof vest a clam uses to give itself a bit more powerful armor. Because pearls are made inside the shell, that’s why the most valuable pearls to us are made by oysters with a nacreous inner shell layer. The tropical pearl oysters (genus Pinctada) make some of the most valuable pearls, because it’s particularly rich in organic material and thus has a bright and complex lustre. Pearl oysters have extremely thin shells, which are strengthened by having a nacreous structure.
Why do bivalves make pearls? Pearls are essentially part of the bivalve’s immune system. If a piece of sand or debris got under your skin, your body would encase the intrusive object with scar tissue. Bivalves can do better than that, because they use the material of the shell to wrap around the object. Anything can be an intrusive object- a piece of sand, an infection, or even a parasite. For example, pearlfish are parasitic fish specialized to live inside the shells of clams. If they die in the shell, the clam will dutifully set to work encasing the fish in nacre, like Han Solo in carbonite!
A pearl oyster with a pearlfish wrapped in nacre against the inner shell. From the Natural History Museum London collection
Most natural pearls are irregularly shaped, so cultured pearls often use round beads as the nucleus for the oyster to grow nacre around. The farmer wedges the oyster’s shell open, deposits the bead and leaves it for a period of time to allow the bivalve to deposit nacre. At harvest time, the oyster can be shucked to remove the pearl. Some experimental approaches even anaesthetize the oyster to remove the pearl, replace it with a new nucleus and repeat the process, potentially allowing for greater efficiency and humane harvesting, allowing the oyster stock to live for years!
So next time you see a pearl, you can understand that the craftsmanship of these wondrous objects is the result of millions of years of evolution, combined with thousands of years of human ingenuity. Moving forward, researchers are attempting to learn to imitate the structure and methods that clams use to make pearls, which could lead to all sorts of improvements in materials science! Clams again prove their skill in engineering. They have a 500 million year head start against us, but we can always learn!
Two giant clams near Eilat in the Northern Red Sea. To the left is the small giant clam, Tridacna maxima, and to the right is a mature individual of the rare endemic giant clam Tridacna squamosina, only found in the Northern Red Sea.
You are what you eat, and clams are too. We’re made of atoms, which come in “flavors” called isotopes, relating back to the mass of the atoms themselves (how many protons and neutrons they have). Nitrogen, for example, comes in two stable (non-radioactive) forms called nitrogen-14 and nitrogen-15. Much like scientists can track the composition of a person’s diet from the isotopes of their hair, researchers have used the isotopes of clams to figure out their diet.
Nitrogen isotopes provide us with a useful and detailed record of food webs. Plants and algae tend to have more of the light isotope of nitrogen in their tissues than the animals that eat them (primary consumers), and the animals that eat those animals have even higher nitrogen isotope values. We can measure the amount of “heavy” atoms of nitrogen with a unit called δ¹⁵N (“delta 15 N”). A carnivore at the top of the food chain will have a very high δ¹⁵N, while plants will be the lowest. Clams, typically being filter feeders, will usually have an intermediate value, since they’re eating a lot of phytoplankton (tiny microscopic floating algae) and zooplankton (animal plankton that eat other plankton).
But I study a special kind of clam, the giant clams, which have a cheat code enabling them to become giant: they have algae *inside* of their bodies. The algae make food using photosynthesis and share it with their hosts! In exchange, the clams provide the algae with a stable environment free of predators, plenty of fertilizer in the form of their own waste, and even channel extra light to help the symbionts grow faster. This partnership is called photosymbiosis, and is pretty rare in clams, though it is common in other animals like the corals that build the reefs where giant clams are found! Previous researchers have shown that giant clams have very low nitrogen isotopic values in their tissue, like a plant, because they get most of their nutrition from the algae, rather than filter feeding.
I am a sclerochronologist. That means I study the hard parts of animals, in this case the shells of bivalves. Like the rings of tree, bivalves make growth lines in their shells which can serve as a diary of their lives. Some of my past work has looked at using chemistry of the growth lines of giant clams to measure the temperatures they grow at, compare the growth of ancient and modern clams, and even look at how much the clams grow in a day! Today though, I’m talking about my most recent paper, which looks at how we can use the shells of giant clams as a food diary.
But when they’re babies, the symbiosis in giant clams is not yet fully developed. During this early period of their lives, giant clams actually get more of their nutrition from filter-feeding like a “normal” non-photosymbiotic clam, until they’ve had a chance to grow in surface area and become a living solar panel. Like all bivalves, the shells of giant clams are made of calcium carbonate, bound together by a protein scaffold we call the shell organic matrix. Proteins are made of amino acids, which contain nitrogen! If we can get the nitrogen out of the shell from the early part of the clam’s life, and compare it to the nitrogen at the end of the clam’s life, it might record the clam’s transition from filter feeding to its mature plant-like lifestyle! If our hypothesis holds, we should record a decrease through its life in the shell δ¹⁵N values.
A model I made of the clams’ nitrogen intake, with the left plot how they switch from filter feeding to getting most of their nitrogen from dissolved sources around 5-6 years of age. Because the nitrogen isotopes of those two sources are different, that manifests in the expected values from the clam’s body (the right plot)!
A map made by my talented partner, Dana Shultz!
So I gathered a team of talented collaborators and set out to test that hypothesis, using giant clam shells that I was able to get on loan from the Hebrew University of Jerusalem Museum. These shells had been confiscated from poachers at the Egypt-Israel border. While I would have rather known these clams were still alive in the waters of the Northern Red Sea, being able to use them for research to understand the biology of their species was the next best thing! I had originally planned on pursuing a postdoc undertaking this project with Rowan Martindale, a professor at UT Austin who has studied the nitrogen isotopes of photosymbiotic corals, but when I started up at Biosphere 2, we ended up continuing with the project anyway as a collaboration! We measured the nitrogen isotopes of the shell material in the lab of Christopher Junium, a professor at Syracuse University, who has developed an exquisitely sensitive method to measure the nitrogen from shell material by essentially burning the shell powder and then scrubbing out unwanted material to isolate the nitrogen, to measure the isotopes in a machine called a mass spectrometer. Katelyn Gray is a specialist in isotopes of biominerals and assisted with drilling out powder from the shells with a Dremel. Shibajyoti Das, now at NOAA, is a specialist in measuring the shell nitrogen isotopes of other bivalves and he was master at doing much of the mass spectrometer work, and assisting in interpretation. Adina Paytan is a professor at UC Santa Cruz. She first provided the funding and support for me to go to the Gulf of Aqaba and collect these shells as part of an NSF-funded student research expedition! She also provided environmental data which helped us to interpret what the clams were actually eating!
A figure showing the four shells we sampled from, with the sampling areas in each hinge area showing colored and matching with the corresponding isotope plot to the right (colored points). 3 of the 4 shells show declines in isotope values with age. Shaded ribbon behind the data shows the model output.
So what did our crack team of scientists find out? We found that three of the four tested giant clams did indeed measure a decline in nitrogen isotopes over the course of their lives. Their earliest growth lines in the hinge areas of their shells record elevated δ¹⁵N values similar to other filter-feeders from the region. But as they aged, their later growth lines show much lower δ¹⁵N values, more like photosymbiotic corals and plants from the region. So clams indeed recorded the transition in nutrition as they became solar-powered! This degree and directionality of change in nitrogen isotopes was much greater than has been observed in any other clams measured in this way, which made sense considering their unique physiology. The clams have another area of the shell, the outer shell layer, which is closer to the symbionts than the hinge area. In this outer shell area, we did not observe much of a consistent trend in nitrogen isotopes. It’s likely that the outer layer is highly influenced by the photosymbionts even at the earliest stages of life.
Growth lines in the hinge area of two of the shells lit from behind, with the drilled areas for this study visible as well. The outer shell layer is the opaque and was also sampled for this study.
There was one clam that differed from the others in showing low δ¹⁵N values through life in its hinge shell layer. To help explain these differences, I created an independent model of the clams’ internal chemistry based on their growth rate, which slows as they age, and also is faster in the summer. When the clams are young filter feeders, they get most of their nitrogen from plankton, debris and other material floating in the water column making up floating material we call Particulate Organic Matter (POM). Meanwhile, when they are in their photosynthetic life stage, they get most of their nitrogen from nitrate, which is essentially Miracle Gro for the symbionts. The model showed that the clams should record a flip from filter feeding to photosynthesis around 4-5 years of age, which was confirmed by three of the shells! But what about the one that didn’t show this trend? My colleague Adina had fortunately measured the isotopes of POM and nitrate in different seasons in the Gulf of Aqaba. We found that in summer, as expected, POM δ¹⁵N is lower than nitrate. In the winter, meanwhile, that relationship is flipped! So if a clam grew more in winter, it would not record the same transition as was seen by the other clams. We think the clam that was the exception to the rule might have been more of a winter grower.
The chaotic nutrient environment of the Northern Red Sea, showing how in different seasons, dissolved nitrate has higher or lower δ¹⁵N values than the Particulate Organic Matter that the clams filter-feed on.
But long story short, we were able to demonstrate for the first time that giant clams show nitrogen isotopic values in their shells in line with expectations from their diet. Other clams have been measured this way, but the fact that we were able to conduct these analyses at all is a testament to the sensitivity of the elemental analyzer in Chris’s lab. Giant clams have *very* low concentrations of organic matter in their shells, so the forward march of technology was a major factor enabling this study to be possible.
Why does it matter that we can measure the transition of the clams from filter-feeding to photosymbiotic in their shell records? Well, giant clams are not the only bivalves which have photosymbionts. There are other clams in the fossil record which have been proposed to have had symbioses with algae, but until now we’ve never had a definitive geochemical way to measure this in fossils. We hope that this approach can be applied to the organic material in fossil shells, which is often well preserved, to see if huge clams in the Cretaceous and Jurassic had a similar way of life to the modern giant clams! If we can demonstrate that was the case, we can see how such species responded to past intervals of climate change, which will help us understand how giant clams will fare in the warming, acidifying ocean of the present.
These results also help explain the lives of giant clams themselves. We hope this kind of data can be used to measure the symbiotic development of giant clams in different places, with different types of food and nitrogen available, where we’d have the potential to measure pollution. Interestingly, the time that the model shows the clams transitioning to photosynthetic maturity is right around the time that they reach reproductive maturity (5-10 years of age). We’d like to investigate whether the time of clam maturity is controlled by the development of their symbiosis, which itself might relate to nutrients in the clams’ environment. If clams can grow faster, then they can mature faster, and potentially reproduce sooner in life. Will giant clams be able to thrive in the presence of increased nitrate, which is a common pollutant in coral reef environments? Like all worthwhile research projects, we have dozens of new questions to pursue as a result of this work, so stay tuned for the next installment in this journey of clam knowledge!
Figure 1 from our paper, showing a comparison of a scallop, its growth increments and where it came from in France, to a giant clam shell section (dyed blue to show its growth lines), and where it came from in the Northern Red Sea
In 2020, I got an interesting email in my inbox from another mollusk researcher! Niels de Winter had emailed me, who I was familiar with from his past work on big Cretaceous rudist bivalves and giant snails. Niels had seen my paper published that year on giant clam shell isotopes from the Gulf of Aqaba in the Northern Red Sea, and was interested in teaming up on a new study to compare the daily growth of giant clams with another bivalve that has daily growth: scallops! I was intrigued because I had similar work underway to study the shells of clams I was growing at Biosphere 2, but I didn’t have any plans to measure my collected wild clam shells that way. So this sounded like a win-win opportunity to work together on a study that neither of us could do alone! Plus, I liked his work and had cited it in the past.
The shells of bivalves are very useful as each produces a shell diary consisting of growth lines, similar to the rings of a tree. Giant clams keep a very detailed diary, with a new growth line forming every night, which previous research has suggested was due to the control that the symbiotic algae inside giant clams have on their host. When the algae conduct photosynthesis, they use CO₂ in the fluid the clam makes its shell from, which increases the pH and accelerates the formation of the shell mineral crystals! The symbionts also directly assist by pumping calcium and other raw materials for the clam to use! Niels had found such daily lines in an ancient rudist bivalve from over 66 million years ago, and proposed it as a sign that the rudists might have had similar algae! I used the daily lines to compare giant clam growth before and after humans arrived in the Red Sea, finding that the clams are growing faster!
But it turns out that giant clams aren’t the only bivalves that make daily lines. Some species of scallops do it too, but that’s a bit confusing, since scallops have no symbionts that could be producing this daily growth period! One way we could investigate this is by bombarding the shells with very tiny laser beams only 20 µm across: the width of a hair is a flawed unit of measurement but 20 microns is as narrow as the narrowest type of hair you can think of! The laser would carry across the cross sections of the shell in a line, literally burning away tiny bits of shell, with the resulting gases captured by a machine called a mass spectrometer, which can figure out the concentrations of elements in the gas.
So we’d basically create a very detailed wiggly graph, where the wiggles represent years, months, days and even tides, depending on how fast the clams and scallops grew! I’m happy to report the paper was published earlier this year, so I thought I’d switch it up a bit and have a conversation with Niels through this blog post. Let me open it up to Niels, who I decided to bring in for this post in a kind of conversation!
Niels, what did you expect to find heading into this experiment? For me, I figured the giant clams would have greater amplitude of variation on a daily basis than the scallops, due to the influence of the symbionts. Is this what you expected? More or less. To be honest, that is what I was hoping to find, because if the daily lines were so much stronger in photosymbiotic shells than in the non-photosymbiotic scallops, it would make it easier to recognize photosymbiosis by studying modern and fossil shells. Also, a finding like that would obviously support the hypothesis we had about the ancient rudist bivalve. However, I was a bit skeptical as to whether the reality would be so clear-cut.
I mailed samples from six juvenile giant clams to Niels for analysis. We went with juveniles for a couple reasons: they grow faster at this life stage than they do as adults: 2-5 centimeters per year for the species we were studying, which meant the greatest opportunity to record a very detailed record from their shells! Scallops also grow extremely quickly, up to 5 cm/year, and so we would be able to get a similar resolution for both types of bivalves, since each page in their diaries would be a similar width.
When I start a new study like this, I always like to “outsource” the expertise about the topic a bit. Our work in sclerochronology often involves bringing together several fields of research and interpreting the results of complex measurements like these requires input from several people who look at them from different viewpoints. I had just finished a research stay at the University of Mainz in 2019, where I worked with Bernd Schöne and Lukas Fröhlich. I know Lukas was working on scallops together with Julien Thébault, whose team collects them alive in the Bay of Brest and keeps a very detailed record of the circumstances the scallops grow at. To carry out the laser measurements, I needed geochemistry experts, and Lennart de Nooijer, Wim Boer and Gert-Jan Reichart came to mind because I was already working with them on other topics and they run a very good lab for these analyses at the Royal Netherlands Institute for Sea Research (NIOZ). This is how the team came together.
Niels conducted a series of laser transects across the clam shells. He used some sophisticated time series analysis approaches to try to quantify the different periodic cycles that appeared in the clam and scallop growth. This was a different approach to how other workers have gone about finding daily growth cycles in giant clams and scallops, where they have often started by zooming in to find the wiggles, and work backwards from there. Niels instead tried to agnostically dissemble the growth records across each clam shell using mathematical approaches, based on the idea that this would be how future workers have to go about identifying daily growth patterns in fossil clams, where we often don’t have a real “growth model” up front to work with. By growth model, I mean the way that we convert the geochemical observations, which are arranged by distance along the shell, into units of time, which requires us to know how fast the clams grew. For the scallops, the age model was made by counting daily “striae” they form on the outside of their shells. For the giant clams, I helped with this by counting tiny growth lines inside the shell made visible by applying a dye called Mutvei’s solution. Because the growth lines weren’t visible all the way through the shell, I used a von Bertalanffy model to bridge across and create a continuous estimate of how old the clams were at each point along their shells.
Niels found some interesting results! I personally expected that the daily variation in giant clams would dwarf what was seen in the scallops, because of the impact of the daily activity of the symbionts. But it turned out that while the clams had a more regular pattern of daily shell growth than the scallops, likely controlled by the symbionts, that was still a minority of the variance across the clams’ records. Yet again, these clams destroyed my hypothesis, but in an interesting way!
Niels, what were your expectations going into this, and how did the results confirm or go against your hypotheses? What challenges did you run into in the course of your analysis, and how did you end up addressing those challenges?
This was honestly one of the most difficult shell-datasets I have worked with so far. The laser technique we used measures the elemental composition of the shells in very high detail, but while this is ideal for funding daily rhythms, it is both a blessing and a curse! In a dataset like this it becomes quite hard to separate the signal we are interested in from the noise that occurs due to measurement uncertainty. I ended up using a technique called spectral analysis, which is often used to detect rhythmic changes in successions of rocks. I guess this is where my geology background was helpful. With this technique, we were able to “filter out” the variability in the records of shell composition that happened at the scale of days and tides and remove the noise and the longer timescale variations. It turns out that, when you do this, you have to remove a surprisingly large fraction of the data, which shows us that the influence of the daily cycle on the composition of both the scallops and the clams is not very large (at most 20%). We did find a larger contribution in the giant clams, as expected, but the difference was much smaller than anticipated. I also find it interesting that most of the variability was not rhythmic. This shows that there are likely processes at play that control the composition of shells on a daily basis which we do not understand yet.
We were measuring a suite of different elements across both bivalve species, including strontium, magnesium, manganese and barium. All of these were reported relative to calcium, the dominant metal ion in the shell material (they’re made of calcium carbonate). This is why we call them “trace” elements; each is integrated into the material of the shell due to a variety of causes, including the temperature, the composition of the seawater, the growth rate of the clams, and also simply due to chance.
Examples of the time series of trace elements from a scallop shell (to the left) and giant clam (to the right), showing the very intricate wiggles in trace element values on a on a tidal and daily basis in each bivalve
In the giant clams, the elements that varied most on a daily basis were strontium and barium. Prior workers had found strontium was the strongest in terms of daily variation, but barium was more unexpected! Normally, barium is thought of as a record of the activity of plankton in the environment, and since there is very little plankton to be found in the Red Sea, it was not expected to see that element vary on a daily basis. It could be that barium gets included in the shell more as a function of the growth rate of the animals. Meanwhile, the scallops (from the Bay of Brest in France) were measuring strong tidal variability in barium and strontium, which makes sense because that location has huge tides compared to the Red Sea. Tides happen on periods of ~12.4 and 24.8 hours. The scallops showed swings lining up with both, and the tidal variability might be the main explanation for how scallops form daily lines. Because the lunar day is so close to a solar day, they would be hard to tell apart from each other! Interestingly, the giant clams also showed some sign of a ~12 hour cycle. While the Red Sea has pretty tiny tides, I had noticed that some of the clams make 2 growth lines a day, and if some clams in the shallowest waters were exposed on a tidal basis, that could explain why they’d make 2 lines: one at low tide, and one at night! Even in places without tides, like the Biosphere 2 ocean, I’d noticed evidence of 12-hour patterns of activity in the clams. It’s so nice (and rare!) when one of my hypotheses is confirmed!
A nice schematic Niels put together showing all the environmental factors that influence the shells of scallops and giant clams, and how much different elements vary as a function of sunlight, tides and other more irregular events like storms. Mn stands for manganese, Ba for barium, Sr for strontium and Mg for magnesium.
Both the giant clams and scallops recorded large irregular swings in all of the studied elements, likely due to non-periodic disturbances. In the case of the scallops, these included storms and the floods of sediment from rivers. For the giant clams, these probably included algae blooms that affect the Red Sea, as well as potentially dust storms that also come every 1-2 years. Both giant clams and scallops have a lot of potential to measure paleo-weather, which is something that other researchers have observed as well!
Niels, where do you see this work heading next?
The recent work looking at very short-term changes in shells is very promising, I think. I agree that there might be a possibility to detect weather patterns in these shells, but that would require some more work into understanding how these animals respond to changes in their environment on an hourly scale and what that response does to their shell composition.
In the meantime, I was intrigued to find that we were not the only people looking for daily cycles in the chemistry of giant clam shells. I had the pleasure of reviewing this paper by Iris Arndt and her colleagues from the university of Frankfurt (Germany). Iris took a similar approach to detecting these daily cycles by using spectral analysis, but she a smart tool called a “wavelet analysis” to visualize the presence of daily rhythms in the shell, which I think was more successful than my approach. She even wrote a small piece of software which can be used to (almost) automatically detect the days and “date” the clam shell based on them. This is quite a step forward, and if I were to do a project like this again, I would certainly try our Iris’ method.
Interesting, too, is that the fossil giant clams studied by Iris showed the daily cycles in magnesium concentration instead of strontium and barium. This shows that the incorporation of trace metals into clam shells is still not fully understood. So one of the things to do, in my opinion, would be to try to see if we can use shells grown under controlled conditions to link the shell composition to short-term changes in the environment. This would require a complex experimental setup in which we simulate an artificial day and night rhythm or an artificial “storm”, but I think it can be done using the culture experiments we do at the NIOZ.
This study represented a unique opportunity to collaborate with my colleague Niels on a topic that interested both of us, which we wouldn’t have been able to pursue on our own. I enjoyed collaborating with him on this work and we have some ideas for further studies down the road, so stay tuned for the next co-clam-boration!
Every clam is a door into the sea. If the “door” of its shell is open, the clam may be happily breathing, or eating, or doing other weirder things. If the door is closed, it may be hiding from a predator, or preventing itself from drying out at low tide, or protecting itself from some other source of stress. It turns out that by monitoring the opening and closing of a clam’s shell valves, a field called valvometry, scientists can learn a lot about the clam’s physiology, its ecology and the environment around it.
Valvometry involves attaching waterproof sensors to each shell valve of the bivalve, to measure the distance between them and their movement. Researchers have used valvometers to figure out that bivalves can be disturbed by underwater pollution like oil spills, harmful algal blooms, and more unexpected sources such as noise and light pollution.
A great video from Tom Scott discussing a Polish program to monitor water quality with valvometry
Giant clams are a group of unusually large bivalves (some species reach up to 3 feet long!) native to coral reefs of the Indo-Pacific, from Australia to Israel. They grow to such large size with the help of symbiotic algae living in their flesh, the same kind that corals partner with the corals that build the reefs. The algae photosynthesize and share the sugars they make with their host clam, and the clam gives the algae nitrogen fertilizer and other nutrients, a safe home from predation and even helps channel light to the algae using reflective cells called iridophores.
A Tridacna derasa clam in the Biosphere ocean. It has deep green flesh, covered with yellow stripes of iridophores and a blue fringe at the edge.
Previous studies have used valvometers on giant clams, but I was always perplexed by how few studies there were: only two that I know of! One study on clams in New Caledonia figured out that the clams partially close every night and bask wide open during the day. The clams’ shell opening behavior and growth was found to become more erratic at temperatures above 27 °C, and when light levels become too great. Another study showed the clams start to clam up when exposed to UV light to protect themselves from a sort of sunburn, which is a real threat in the shallow reef waters they live in.
Two clams sitting next to each other in the B2 ocean. They often moved themselves to “snuggle” next to each other this way. Safety in numbers!
There is clearly a lot of information to pick up about how clams react to their environments, which can help us understand the health of the clams and also the corals around them. Coral reefs are under global stress from climate change, overfishing and pollution. Giant clams are some of the most prolific and widespread bivalve inhabitants of reefs, and represent an appealing potential biomonitor of reef conditions. Many giant clam species are threatened by the same stressors that influence the corals which build the reefs they live on, as well as overharvesting for food and their shells. For that reason, wild examples should clearly not be bothered by applying valvometric sensors. But giant clams are increasingly grown for the aquarium trade, resulting in a wealth of cultured specimens which could serve as sentinels of reef health, if they were fitted out with sensors. All of these motivators made me more and more curious of why we don’t have more literature monitoring the behavior of these clams with valve sensors.
I wondered if one of the limiting factors preventing the use of valvometry on giant clams is expense and ease of access. Giant clams live primarily in regions bordering developing countries in the Indo-Pacific, and almost all the professional aquaculture of clams for the reef trade happens in such countries, including places like Palau, Thailand, and New Caledonia. These countries are far removed from the places where most of the proprietary valvometric systems are manufactured. These systems can cost several thousand dollars even in Europe, never mind Palau, where arranging the import of electronics can be difficult.
When I started my postdoctoral fellowship at Biosphere 2 in 2020, I set out to grow two dozen smooth giant clams (Tridacna derasa, a species which can grow to about 2 feet long) in the controlled environment of the Biosphere 2 ocean, a 700,000 gallon (over 2.6 million liter) saltwater tank used to grow corals and tropical fish and kept at a stable year-round temperature of 25 °C. We suspended a series of LED lights intended to simulate the powerful light levels these clams experience in the wild (light is a lot brighter in the tropics than it is in Arizona!). The main focus of my project involved measuring the shell chemistry of the clams, to determine how their body chemistry changed as they grew from mostly getting their energy from filtering algae food from the water like other clams, to getting most of their energy from sunlight like a plant. But as a “side project” I set about measuring the behavior of the clams with custom-built valvometers based on open-source, inexpensive hardware that would be more accessible to researchers in the developing world. That work has since been published in PLoS One!
In our design, we used Hall effect sensors. Hall effect sensors generate a voltage when a change in magnetic field is detected. They are cheap, easily obtained for less than $1.50 apiece and are common in the electronics hobby trade. You might have encountered one in a home security system door/window sensor, where they help detect if a door is open or shut. We stuck a hall sensor soldered to a long copper cable to one valve of a clam, and a small magnet to the other valve. When the clam closed, we could measure exactly how closed it was. You can see why I started off by calling clams doors into the sea: we were literally measuring them that way!
Showing the sensor soldered to the three strands of the cable.
But here the first challenge of my project appeared. The off-the-shelf Hall sensors don’t come in waterproof form, and I learned quickly that the ocean really, really loves to break my gear. After dozens of failures, I settled on coating the sensors in waterproof grease, wrapping that in heat-shrink tubing and then sealing that inside of aquarium-grade silicone. During this process, a gifted technician at Biosphere 2 named Douglas Cline helped with iterating on the first prototypes. At a certain point I taught myself to solder so I could do my part to improve the sensors.
It was also hard to figure out how to attach the sensors and magnets to the clams in a durable way. Neither of the prior studies mentioned how they attached the sensors to giant clams, and I tried and failed with literally a dozen different ways before settling on “pool putty,” a two-part adhesive often used to seal leaks in pools that can cure underwater. I found the pool putty had trouble attaching to the clams’ shells on its own, so I combined it with a special kind of cyanoacrylate superglue called “frag glue,” often used to attach pieces of corals to growth stubs. I also had to find a way to attach it to the clams without stressing them out. I determined five minutes out of the water was enough time to get the sensors attached to the clams, after which they could be returned to the water to finish curing. While giant clams are adapted to spend extended periods out of the water in their natural intertidal environment, we wanted to make sure to minimize their stress however possible, to ensure they would show natural cycles of behavior in the data.
Figure from the paper showing: A) schematic of the sensor attached to the clam, linked to an Arduino microcontroller and Raspberry pi computer. B) A sensor attached to one of the clams
We were pleased to see the cyborg clams seemed to pay no mind to the sensors. Giant clams are adapted to encourage all sorts of other critters to live on their shells as a form of natural camouflage, and I think the clams interpreted the sensors as pieces of coral or anemones sticking to the side of their shell. Whatever the case, as long as we kept the cable pointing to the side away from the clams’ flesh, they opened five minutes after being returned to the water, and their behavior and growth rates were indistinguishable from the clams that didn’t have sensors attached.
One of what became many sunsets on the Biosphere 2 ocean shore troubleshooting the clam sensors! Pardon the chaos: mad scientist at work!
So how did we measure the voltages coming from the sensors? Our design featured an Arduino microcontroller, sort of like a smart circuit board which can measure the voltages coming back over the copper cables. Arduinos are very cheap, and we chose a $25 model. Even more importantly, Arduino has a huge library of plug-ins available to keep the exact time of each observation using a clock attachment, and the data can be uploaded to SD cards or an attached computer. For the attached computer, I used a Raspberry Pi computer, which are open-source Linux-based tiny computers that are very cheap! Or rather they were very cheap before the pandemic, but fortunately there a lot of open-source alternatives that can be obtained more cheaply. We logged the data on the Raspberry Pi as it rolled over from the Arduino, and I could watch the read-out on a monitor right on the Biosphere 2 beach. We set the Arduino to record every 5 seconds.
Sensors attached to four of the clams. Notice the one on top left has closed a bit, after sensing my presence! They have eyes so they were able to detect me 😀
We ran the sensors for three months. During that time, the baby giant clams grew almost an inch! What did the sensors record them doing? During the day, the clams basked wide-open, exposing as much of their tissue as possible to light (other than the times that I disturbed them by swimming above them, of course)! This schedule of opening aligned pretty closely with the times that maximum sunlight hit their part of the Biosphere 2 ocean: the mornings, because the clams were on the east side of the building. At this time of day, the clams want to expose their symbiotic algae to as much light as possible, so they can conduct photosynthesis and make sugars that the clams use as food!
A) Plot of the valvometry data. Points higher on the plot mean the clam was more closed, up to 100% closed. The clams proceeded by opening in the early morning and then closing in the early afternoon. The big red circles represent times that the clams closed briefly, with bigger circles representing a longer time spent closed. Most of these rapid closures happened at night. B. A plot of Photosynthetically active Radiation (the amount of light the clams had to use for photosynthesis). The highest values were in the mid-morning when the clam lights were running in combination with direct sunlight hitting them from above.
Around mid-afternoon, the clams started to close partially, to about half closed. Why might that be? My hypothesis is that this posture represents a kind of “defensive crouch” to protect themselves from predators, in this case fireworms that live in the Biosphere 2 Ocean and were constantly kicking the clams’ tires. Similar nighttime behavior was observed in wild clams in a previous study, but not in a study that took place in a small predator-free terrarium tank. By remaining partially closed, the clams are prepared to rapidly close completely if they feel a predator approaching. But they only expend that energy of staying in that posture if predators are around!
One of the fireworms that proved to be my nemesis and continually attacked the clams during the experiment
And approach the fireworms did. We observed frequent closures at night lasting anywhere from a few seconds to hours, likely partially related to the activity of the worms around the clams. But the clams were engaging in another activity at night: filter feeding! Giant clams really get to have their cake and eat it too, because during the day, they act like a plant, but at night, they eat other plants in the form of plankton that they filter feed out of the water using their gills! At regular intervals, the clams need to clear uneaten material from their gills in a process sometimes called “valve-clapping”. The clams yank their shell valves together rapidly to force water out, blowing out pseudofeces: unwanted material packaged with mucus. We measured this valve-clapping mostly at night. The clams are likely scheduling this activity for the night-time so they can prioritize staying open and filter feeding during the day!
Figure comparing how often clams closed per day to measures of how high plankton numbers were in the Biosphere 2 ocean (chlorophyll is a marker of phytoplankton while phycocyanin is a measure of cyanobacteria), and how high the light levels were. Peaks in closure activity often happened shortly after rises in algae.
We observed that the frequency of valve clapping aligned closely with the rises and falls of chlorophyll concentration in the Biosphere 2 ocean, which is a measure of how much plankton is in the water column. The clams would engage in a burst of valve clapping around 4 days on average after a bloom in chlorophyll, suggesting they were filtering out plankton after they had died and settled to the bottom where the clams could eat them. We also found that the clam’s filtering activity peaked at times of highest pH. This likely is due to the fact that higher pH means the algae around the clams are being more active, and pulling CO2 in from the water to use in photosynthesis, making the water less acidic. More photosynthesis means potentially more material for the clams to filter through! This data helps quantify how giant clams help filter the water in their native environments! Coral reefs depend on very clear transparent water to allow maximum sunlight to reach the corals, and the filtering activity of giant clams likely plays a big role in helping preserve those conditions!
So we found that by adding sensors to clams, we could record their ability to feed from the sun, their feeding on plankton around them and their avoidance of predators. How can this technique be used next? We hope that by using cheap off-the-shelf resources and open-source software, we can enable more sensors to be put on clams all over the world, such as places where giant clams are farmed in Palau, New Caledonia, Thailand, Taiwan, Malaysia and more! If we can collect data on clam activity from all these places, we can compare how their feeding patterns differ in places that have more or less plankton floating by, or have more or less sunlight available, or different predators that affect the clams’ behavior. This data would have importance to the clams’ conservation, as well as our understanding of the reef overall. In future years, I hope we can develop a global network of cyborg giant clams from the Red Sea to the Great Barrier Reef, so we can better understand how these oversized and conspicuous but still mysterious bivalve work their magic!
Abraclam Lincoln (photo source) next to a handful of Mercenaria mercenaria, a closely related kind of quahog (photo from Wikipedia), illustrating that Abraclam is indeed a chonker. To the right we have Arctica islandica, the famous long-lived clam, illustrating how hard it is to tell these clams apart without specialized training (photo from conchology.be).
Recently, a bit of an amusing clamfuffel arose when the Gulf Specimen Marine Lab, a research institute in Florida, began posting about a supposedly 214-year-old clam they named “Abraclam Lincoln”, in honor of potentially sharing a birth year with Honest Abe. The story went viral, and while some of their clammy claims turned out to hinge on flawed assumptions about how clams can be aged, it was still a worthwhile opportunity to communicate about the wonderful world of bivalves with people.
I was particularly impressed with how GSML went out of their way to correct the record as more info came in. Long story short, Abraclam is not the long-lived ocean quahog Arctica islandica (more on them below), but in fact a specimen of the southern quahog Mercenaria campechiensis, which lives along the Gulf coast and grows much more quickly than A. islandica. So the large clam they found was likely more like 40 years old, which is not too unusual for this species, rather than two centuries old. It also makes much more sense to find Mercenaria in Florida than Arctica. Additionally, external shell growth lines in Mercenaria are known to not be reliable for aging the species. The shell would have to be cut open to view internal lines to figure out how old Abraclam really is, which would require killing them. Fortunately, they released the clam rather than cut them in half for science.
Known range of Arctica islandica as a heatmap, compared to location of GSML on the Florida Panhandle. Source: OBIS
Slice through a Mercenaria campechiensis shell. The shell is lit from behind to show the annual growth lines. From Moss et al. 2021, written by fellow clam man David Moss, a friend of mine!
For what it’s worth, I still think Abraclam is an interesting specimen of M. campechiensis– they have a huge scar in their shell that would be interesting to learn more about (maybe they were hit by a dredge in their youth, healed the break and recovered to reach a ripe old age), and an unusual undulating margin at the edge of the shell that could be a deformity reported in the past literature for this species. Sometimes, scientists are wrong the first time, but we’re open about how we’re wrong, and everyone ends up learning more than we would have known otherwise.
So far so good. Then a bull had to wander into this delicate china shell with the entry of Stephen Colbert into the debate. I’ll let Stephen speak for himself, but needless to say, after his rant I feel a need to respond:
Before I dig into this clambake, Stephen, kudos to you for covering the whole story and not just the initial incorrect information. You addressed all the big inaccuracies, from the size not being tremendously out of the ordinary, to the incorrect species ID, to the incorrect age. Maybe it’s not so bad that people are getting their news from comedians rather than the news media these days! But while there were definitely some pearls of wisdom within your monologue, I have to point out some misconceptions here.
“The only thing more heartbreaking than the lies we were fed in this story…is growing up to be a clam expert!”
– Stephen Colbert
This is just plain false, since I’m not heartbroken, because clams are frickin’ awesome. Clams are way cooler than you or me, and that means by extension that the people who study them are pretty cool and interesting too (not really referring to myself. I’m just an eclamgelist along for the ride). So here are three facts about big old clams, and information about the clam experts who discovered these facts!
The ocean quahog lives to >500 years old!
Arctica islandica shell I saw on the beach in Massachusetts. This individual was likely several decades old when it died based on its fairly large size!
The ocean quahog Arctica islandica (which Abraclam was initially misidentified as) is tremendously long-lived, one of the longest-lived animals on earth! It has been confirmed to live to at least half a millenium! One individual caught off the coast of Iceland was aged to ~507 years by counting tiny growth lines in its shell via microscope, combined with radiocarbon dating. This clam was named Hafrún (meaning “ocean mystery” in Icelandic), but is sometimes called Ming due to it being born in the Ming dynasty of China. So put that in your Ming vase and smoke it Colbert!
Part of the shell of Hafrún, which was cut open to determine its age via internal growth lines. Source: Museum Wales
Several scientists worked on aging Ming, but Alan Wanamaker at Iowa State was a lead author on the original work. He uses growth lines in the shells of many clam species as records of climate change and is generally one of the nicest people you could have the opportunity to meet.
Tridacna gigas grows to over 4 feet long and hundreds of pounds!
Large specimens of giant clams that are around 3 feet long at the California Academy of Science collection.
The author sitting in a scale model of Tridacna gigas at the Monterey Bay Aquarium
Abraclam might be only 6 inches long (which is respectable as he/she is quite girthy; length isn’t everything, Stephen!), but there are other types of clams that are bigger than one Colbert in mass. The giant clam Tridacna gigas grows to over 4 feet long and weighs hundreds of pounds. They live on tropical coral reefs and use the power of the photosynthetic algae in their flesh to speed up their growth. So basically these clams are bigger and way more interesting than you, Stephen, since they get to go out and tan in the sun for lunch while you have to gobble down a slice of pizza.
Mei-Lin Neo at University of Singapore is considered the world’s leading expert on Tridacna, and has done more than almost anyone I know to describe all twelve currently known species of giant clams found around the world. She’s a tremendous advocate for giant clam conservation and gave an outstanding TED talk about them to boot. You should have her on your show to be honest.
Geoducks: they looked like that first!
A 6.5 pound geoduck and admiring Washington Department of Fish and Wildlife Volunteer (Source)
Speaking of girthy, long clams, I’d be remiss not to mention the geoduck, Panopea generosa. Pronounced “gooey-duck,” these clams looked like this long before any part of human anatomy existed, having been around in various forms since at least the Jurassic. They have a long siphon that they use like a snorkel when they dig deep in the mud, and they can live for almost 200 years.
Brian Black at the University of Arizona is an expert in using their shells as a record of climate change. He was part of a group that was able to stitch together the growth line records from multiple geoduck shells to make a continuous record of climate change going back to 1725. Seems appropriate to note that 1725 was the year that Casanova was born…a man who may have channeled some qualities of geoducks.
Local experts on Abraclam
I’d like to mention two of the experts who corrected the record about Abraclam Lincoln and provoked Stephen’s attack in the first place. Dr. Dan Marelli wrote an op-ed correcting the record on how Mercenaria clams are aged for the Tallahassee Democrat. He’s an expert scientific diver and has published papers on clams ranging from endangered scallops to invasive mussels. Scientific diving is crucial to understand clams in their native environment, and to assist in their conservation. If I had to choose who had more interesting stories at the bar, it’d be an easy decision to listen to the swashbuckling diver over the late-night TV host!
Dr. Edward Petuch at Florida Atlantic University reached out to GSML to make sure they knew the correct species ID for Abraclam. He is well-known for his work describing the change in ecology of mollusks in Florida and the Caribbean over the last several million years. GSML expressed interest in working with Dr. Petuch in the future, and I can confirm that I’ve had fruitful scientific collaborations start when other scientists have reached out to me about how I was totally, embarrassingly wrong. Being wrong in science is part of the job, and that’s why I’m glad this Abraclam story came out in the first place.
“So what does your son do? He’s a marine biologist. Does he work with dolphins? …I’m gonna say yes.”
-Stephen Colbert
To close out, I’d like to address Stephen’s assertion that my mom isn’t proud of me for being a clam expert. Stephen, I’ll have you know that my mom is the most enthusiastic patron of my clam science. She reached out to the local paper to anonymously tip them to interview me about my clam work, had me give a speech about clams at the local women’s group she’s a part of, and when I defended my PhD thesis, she made t-shirts to commemorate the occasion. I can confidently say I wouldn’t be Dan the Clam Man if it weren’t for her support. Thanks Mom!
The tree of life is often portrayed as a neatly branching structure, with each division point cleanly delineated and separated from its neighbors. The truth is that the various twigs of the tree of life often overlap and become tangled in a process we call symbiosis. I’ve talked about symbiosis before on this blog, which falls along a spectrum of wholesomeness. At one end we have mutualism, a partnership where both organisms benefit and achieve more than the sum of their parts. The other extreme is parasitism, when one organism benefits at the expense of the other. Between the two, there is a broad gray area including commensalism, when one organism’s presence doesn’t necessarily cost or benefit the other in any way. The tree of life is crowded and unpruned, and so sometimes the twigs might wrap around each other quietly and without much fuss. We live on a small planet, and have had to get used to living in uncomfortable intimacy with all sorts of creatures, such as the mites that are living on your eyelashes right now.
But things start to get really weird and tangled when the tree of life loops over on itself twice, or three times, or more. “Three-way” symbioses are surprisingly common, and the more you look for them, the more you realize that the tree of life is more of a knot than anything else.
A view of the Heteropsammia coral from the side
A recent paper from researchers in Bremen (Germany) and Saudi Arabia looked at such a three-way symbiosis between a coral, a worm and bivalves found off of Tanzania in East Africa. The relationship between solitary corals (Heteropsammia cochlea and Heterocyathus aequicostatus), a sipunculan worm (Aspidosiphon muelleri muelleri) and the clam Jousseaumiella, is a complex triangle of dependencies that had previously been noticed by other researchers, but never investigated at great depth. The worm lives with multiple tiny clams attached, all inside of a small solitary coral the size of a dime (1 cm long). Is the coral a willing host for this crowded boarding house, or has it been parasitized? Does the worm gain anything from the clams? The researchers sought to find out.
Part of the reason I enjoyed reading this study so much was that it had to take a narrative structure to describe the evolutionary ménage à trois of its focus. So much of modern science has moved away from anecdote to hard data, and while there is plenty of that to find in the study, it turns out that a lot of the study of symbiosis is storytelling. We need to know the setting and the characters.
In this case, the main characters are small solitary corals living in the tropical reefs of the Indo-Pacific. We denote them as solitary to distinguish them from their giant colony-forming compatriots that construct the coral reefs currently threatened by climate change and pollution. But like those giant reef-builders, these solitary corals get much of their food from sunlight through a mutualistic partnership with algae called Symbiodinium. The algae provide the host with sugars and other photosynthetic products, and the hosts give them nutrients and a safe cozy home in their tissue.
You might be thinking, “Wait! Dan just said this was a three-way partnership between a coral, a worm and some clams. So this is actually a four-way partnership between corals, worms, clams and algae?” You’d be exactly right. And I’m happy to say that the plot of this sordid story is about to thicken even further.
The side of the coral. See the little pores?
The Aspidosiphon worm is found in a spiral-shaped burrow inside of the skeleton of the coral. It is a pretty cozy home, with walls made of calcium carbonate by the coral, with breathing holes in the sides to allow the worm to breathe and release waste. The researchers wanted to know more about the structure of the burrow. Was it dug out by the worm using acid or an abrasive motion, like some clams use to dig into coral? So the researchers essentially gave the coral a CT scan to see its 3D internal structure. Inside they found growth features suggesting the coral grew around the worm, as if intentionally providing it a home.
Cross-sectional CT scans of the coral skeleton. In figure D, you can see the silhouette of the chambers of the snail shell where the worm made its first home!
Even more crazily, they found evidence that the worm had first settled inside an empty snail shell, like a hermit crab! The coral probably settled on a snail shell as a larva, and grew to engulf the whole snail shell, leaving growing space for the worm inside, with windows and all! So to review, this is now a five-way symbiosis between a dead snail, a worm that moved into its empty shell, the coral (powered by algae) that grew around it and encased the snail shell within its skeleton, and we haven’t even gotten to the clams. How many creatures are hiding stacked in this trench coat? Please bear with me as I explain!
An SEM image of Jousseaumiella. These are less than 1 mm long! Pinhead sized!
What are the clams doing in this picture? Jousseaumiella is part of a family of clams called Galeommatidae, which we previously mentioned on this blog in the context of some bivalves found growing in the gills of unfortunate sand crabs. Many members of the Galeommatidae family are parasitic or commensal with other marine organisms. In this case, Jousseaumiella are tiny flat-bodied clams less than 1 mm long, found attached to the body of the worm, squeezed inside the burrow in the coral’s skeleton. It feeds on the worm’s waste and potentially food particles coming through the pores in the sides of the burrow. Not the most dignified existence, but a more mobile home means more opportunities to eat a varied diet similar to that that the worm and coral are seeking out, and the clam also gets protection from predation tucked inside the coral. It is unclear if it benefits the worm directly to have clams attached to it.
A time lapse of the coral+worm moving from the paper’s supplement! It would be handy to navigate to greener pastures, if it became too muddy in a certain place!
It is, however, clear how living inside a coral would be a pretty good deal for the worm, which gets a stable, protective suit of carbonate armor to protect it from predators, and grows to fit it as it gets larger. They are normally found inside of rocks, shells and other hard inanimate objects, but having a living home is a cool upgrade. What is the coral getting out of the deal? The researchers note that the corals are often found in the crevices between other large reef-building corals, in areas of the reef that receive high supplies of nutrients and turbidity (dirt that blocks out light). These sorts of environments aren’t necessarily friendly places for a coral to be, since they reduce the light and therefore the food that the coral can receive from photosynthesis. These crevices also have a lot of variability in other conditions like temperature and water flow. But because the coral has hitched a ride on the back of a worm, it can actually move in the sediment to react to changing conditions and avoid being buried by piles of sediment floating by! The worm can also act as a sort of anchor preventing the worm from sinking in the sediment underneath, which would be a big hazard for the small, stubby coral on its own. The coral seems to go to great pains to make its partner comfortable, not growing its skeleton to cover the pore windows to the outside. The researchers note that as coral reefs worldwide are subject to increasing human-made pollution and climate change, it would be interesting to research whether this complex three-(five?) way symbiosis provides the various participants with an advantage compared to other corals.
So like any good story, this symbiosis features complex, growing characters, a dynamic setting, and still plenty of mystery demanding a sequel! To that end, there are lots of other great three-way symbioses to investigate. Snails which farm fungus that parasitizes plants. Bryozoans living on snail shells that have a hermit crab inside. Gobies serving as lookouts at the entrances of burrows built by shrimp, with a crab freeloader along for good measure. Algae and bacteria teaming up to attack mussels. The list keeps going! I could see this becoming quite a franchise!
Two giant clams off the coast of Israel. Left: Tridacna maxima, the small giant clam. Right: Tridacna squamosina
Another year, another new paper is out, another clamsplainer to write! The fourth chapter from my PhD thesis was just published in Proceedings of the Royal Society B. This study represents five years of work, so it feels great to finally have it leave the nest. In this study, we investigated the comparative growth of fossil and modern giant clams in the Gulf of Aqaba, Northern Red Sea. Back in 2016 during my PhD, I knew I wanted to study giant clams because they are unique “hypercalcifying” bivalves that grow to huge sizes with the help of symbiotic algae living in their bodies. The clams are essentially solar-powered, and use the same type of algae that reef-building corals depend on! Unlike corals, which are the subject of a ton of research related to how they are threatened by climate change, habitat destruction and pollution, comparatively little is known of how giant clams will fare in the face of these environmental changes. Are they more resistant than corals, or more vulnerable?
T. squamosa on the reef off the coast of Eilat.
I had strong reason to suspect that the clams are struggling in the face of human changes to the environment. They can bleach like corals do when exposed to warm water, and have been observed to be harmed when waters are less clear since they are so reliant on bright sunlight to make their food. But I need a way to prove whether that was the case for the Red Sea. I needed to travel to a place where fossil and modern giant clams could be found side by side, so their growth could be compared using sclerochronology. We would count growth lines in their shells to figure out how fast the grandaddy clams grew before humans were around, and compare that ancient baseline to the growth rate of the clams in the present. Giant clams make growth lines every day in their shells, giving us the page numbers in their diary so we can figure out exactly how fast they grew! We can also measure the chemistry of their shells to figure out the temperatures they experienced from the oxygen isotopes, and even what they were eating from the nitrogen isotopes.
A map of the Gulf of Aqaba, where our study took place. My talented marine scientist partner Dana Shultz made this map!
It just so happened that UCSC’s Dr. Adina Paytan was leading an NSF-funded expedition to the Red Sea in summer 2016, which represented a perfect place to do this work. There are many age dated fossil reefs uplifted onto land around the Gulf of Aqaba on the coasts of Israel and Jordan, and there are three species of giant clam living in the Red Sea today: Tridacna maxima (the small giant clam), Tridacna squamosa (the fluted giant clam), and Tridacna squamosina. Tridacna squamosina is particularly special because it is only found in the Red Sea, making it an endemic species. It is extremely rare in the modern day, with likely only dozens of individuals left, making it potentially endangered.
So I set off with Adina and two other students to live for two months in in the blazing hot desert resort town of Eilat, Israel, working at the famous Interuniversity Institute. Getting a permit from the Israeli National Parks Authority, I collected dozens of empty giant clam shells (no clams were harmed in the course of this study!) from the surf zone and from ancient reefs ranging from a few thousand years to almost 180,000 years old. I also spent a week over the border in Aqaba, Jordan where I worked with Dr. Tariq Al-Najjar, my coauthor and director of the University of Jordan Marine Science Station. Tariq is a specialist in algal productivity in the Gulf and was an excellent resource in trying to understand how water quality has changed in the area through time. He pointed out that over the years, the Gulf of Aqaba has had an increased nutrient supply far above what it received in historic times. For nearly 20 years the Gulf was subjected to excess nutrients from Israeli fish farms, which caused tremendous damage to the reefs of the area with their releases of fish waste. The farms were finally forced to close after a long lobbying campaign from Israeli and Jordanian scientists and environmentalists. But even after the farm pollution stopped, there was still increased nutrient supply from runoff and even carried into the Red Sea by dust in the form of nitrate aerosols. These aerosols are produced when our cars and power plants release nitrogen oxide gases, which react in the atmosphere to form nitrate and fall during periodic dust storms that hit the Red Sea a few times per year.
All of these sources of nitrogen are fertilizer for plankton, causing what scientists call “eutrophication.” When plankton blooms, it literally causes the water to be less transparent, which could reduce the clams’ ability to gather light and lead to them growing more slowly. At least that was my hypothesis, but I had to prove if it was true or not. So during that summer and over the next few months, I cut dozens of clam shells into cross-sections, used a special blue dye called Mutvei solution to make their growth lines visible, and took pictures of those lines with a microscope. Then I counted those lines to figure out how many micrometers the clam was growing per day.
A picture showing some of the fine daily lines visible in a blue-stained shell
Here I hit my first challenge: it turned out some clams were putting down one line per day as expected, but some were putting down twice as many! But the way I was using to discern between the two was to measure the oxygen isotopes of the clams’ shells, which forms a record of temperature. By counting how many lines appear between each annual peak of temperature, we confirmed some were daily and some were twice daily. But the oxygen isotope approach is expensive would not be scalable across the dozens of shells I had collected.
Annual growth lines in the shell of a Tridacna maxima clam
Then I remembered that I could measure the lines in the inner part of the clams’ shells, which are formed annually. By counting those lines and then measuring the length of the clam, I could get an approximate measure of how much it grew per year on average. This would allow me to calibrate my band-counting and discern which records represented daily lines and which were twice daily! What a relief.
So I went through all of the shells, counting lines and gathering growth info for as many shells as I could muster. It meant many hours staring at a microscope, taking pictures and stitching the pictures together, then squinting at my computer screen highlighting and measuring the distance between each growth line. I had hoped to come up with an automated way to measure it, but the lines turned out to be faint and difficult for the computer to distinguish in a numerical way. So instead I just powered through manually. When I had the raw growth data, I then transformed them to a pair of growth constants commonly used in the fisheries literature to compare growth across populations. When I put the data together across all 55 shells, I was surprised to discover that my hypothesis was totally incorrect. The clams were growing faster!
Growth constants for all three species, comparing fossils and modern shells. We used two growth constants (phi prime and k) to help control for the fact that our clams were at different sizes from each other. You wouldn’t compare the growth rate of babies and teenagers and try to make any broader assumptions of their relative nutrition without some additional attempts to normalize the data!
Science rarely goes according to plan. The natural world is too complex for us to follow our hunches in understanding it, which is the main reason the scientific method came about! But at a human level, it can still be shocking to realize your data says you were totally wrong. So after a few days sitting and ruminating on these results and what they meant, I remembered what Tariq and other scientists had said about nitrates. The clams are essentially part plant. They use photosynthetic symbionts to gain most of their energy. And much as nitrate pollution can fertilize plankton algae growth, maybe it could do the same for the algae within the clams! It had previously been observed that captive giant clams grew faster when “fertilized” with nitrates or ammonia. But such an effect had never before been observed in the wild. We needed a way to demonstrate whether the Red Sea clams were experiencing this.
Fortunately, the clams also keep a chemical record of what they’re eating within the organic content of their shells. Shells are a biological mineral, made of crystals of a mineral called calcium carbonate. But within and between those crystals, there’s a network of proteins the clam uses like a scaffold to build its shell. Those proteins are made of amino acids that contain nitrogen. That nitrogen comes in different “flavors” called isotopes that can tell us a lot about what an animal eats and how it lives. The ratio of heavier nitrogen-13 and lighter nitrogen-12 increases as you go up the food chain. Plants and other autotrophs have the lowest nitrogen isotope values because they use nitrate directly from the environment. For every level of the animal food chain, nitrogen isotope values increase. Herbivores are lower than carnivores. If you live on only steak, your nitrogen isotope values will be higher than a vegetarian. The same will be true for clams. If the clams were taking in more nitrogen from sources like sewage or fish farms, they would show higher nitrogen isotope values in the modern day.
We found that nitrogen isotope values were lower in the modern day!
Taking bits of powder from several dozen of the shells, we worked with technician Colin Carney at the UCSC Stable Isotope Lab to measure the nitrogen isotopes of the shell material. A machine called an Elemental Analyzer literally burns the shell material to release it in a gas form. A carbon dioxide scrubber absorbs the CO2 and carbon monoxide gas, leaving only the nitrogen gas behind. That gas is measured by a mass spectrometer, which essentially separates out the different isotopes of nitrogen and tells us what fraction is nitrogen-13 or nitrogen-12. Plotting all the shell data together, I discovered that my hypothesis was…totally wrong. The nitrogen isotope values of the modern shells were lower than the fossils. The clams had moved down in the food chain, but how?
A dust storm rolling over the Israeli Negev Desert. Source
After ruling out a bunch of other explanations including the preservation of the shells, we propose that this represents a human-related change in the environment that the clams are recording. As I mentioned before, the Red Sea these days is regularly hit by huge dust storms which are conduits for nitrate aerosols. Our cars emit nitrogen-containing gases which, through a complex web of chemical reactions in the atmosphere, end up in the form of nitrate particles called aerosols. These nitrate aerosols bind to the dust delivered by strong windstorms called haboobs, which carry the dust long distances, with some of it being deposited several times of year. This deposition of nitrate has been found to form up to a third of the nitrate supply hitting the Red Sea, and was a source of nitrogen that wasn’t available to the clams in historic times. These nitrate aerosols are extremely low in nitrogen isotope value, and would be very likely to explain the lower nitrogen isotope value in our clams! If the clams ate the nitrate, their symbionts would grow more quickly, providing them with more sugars through photosynthesis and accelerating clam growth!
Some additional factors probably also have influenced giant clam growth in the region. The Red Sea historically had regular monsoon rains which likely slowed growth in fossil clams, as storms are known to do for giant clams in other areas, but such monsoons no longer reach the area. The Red Sea also had much higher seasonal range of temperatures in the past, with colder winters and warmer summers. Both factors (storms and extremes of temperature) have been previously shown to depress giant clam growth, and so the modern Red Sea may be a goldilocks environment for the clams: a consistent year-round not too cold or hot temperature.
However, as we discuss in the paper, these factors don’t necessarily mean that the clams are healthier. Faster giant clam growth has been found in other research to lead to more disordered microstructure in their shells, which would have uncertain effects on their survival against predators like fish, lobsters and humans. Additionally, a higher nutrient supply to reefs often causes the corals that build the reefs to lose out to competition from algae that block sunlight and crowd out coral colonies. If the reefs are harmed by the climatic changes that have potentially helped the clams, the clams will still lose. Giant clams are adapted to live only where coral reefs are found, and nowhere else. So more research will be needed in the Red Sea to determine if the health of clams and corals is hurt or harmed by these nitrate aerosols, and what that will mean for their long-term survival in the area.
Over the course of working on this research, the giant clams taught me a lot about life. They taught me that my hypotheses are often wrong, but that’s alright, because my hypotheses can still be wrong in a way that is interesting. I learned to go with the flow and trust the clams to tell me their story through the diaries they keep in their shells. I have followed their lessons wherever they led. Now I am doing follow-up work growing giant clams in a giant coral reef tank at Biosphere 2 in Arizona, to directly observe how the clams’ symbiosis develops and create new forms of chemical records of their symbiosis! The work described in my paper here has led to a suite of different ongoing projects. The clams have many more lessons to teach me. Thank you clams!
Right: Gollum, Middle Earth. Photo credit: Peter Jackson, New Line Cinema. Left: Congeria kusceri. Photo credit: Helena Bilandžija
In some ways, living in a cave sounds pretty comfortable. A relatively stable year-round environment and refuge from above-ground predators/competitors make it a place that trogloditic (cave dwelling) animals can develop stable evolutionary forms which persist through long periods of time. Like the deep sea, caves are great places to look for “relict” organisms representative of groups that disappeared long ago on the surface. But living in a cave presents a lot of challenges that require specialized adaptations. The most obvious is the lack of light, the source of energy for photosynthesis supports food webs on the surface. Animals in caves must make a living eating material that flows in from the surface world, like bat gauno or debris from runoff. They can also make a living eating the microbes that directly feed off the rocks of the cave walls. Some caves also have methane or other geologically sourced gases that can serve as a food source, similar to the deep sea, where methane seeps and hydrothermal vents provide fuel for diverse and vibrant ecosystems.
You don’t need to watch Lord of the Rings to see that spending your life in a cave like Gollum requires serious changes to your lifestyle. But that’s exactly where we can find an unusual genus of freshwater mussels called Congeria. Congeria consists of three species found so far only in fifteen caves in the Dinaric Alps, a mountain range in Southeastern Europe stretching from Italy to Albania. These mountains are made from limestone that first formed at the bottom of a shallow tropical sea (picture the Bahamas and you get a good idea of how it would have looked) during the Mesozoic era, the time of the dinosaurs. This limestone (also called karst) was lifted up in the early Cenozoic (after the Cretaceous extinction) by tectonic activity due to the collision of the Adriatic microplate with Europe. This 645 km-long row of mountains is made of a highly soluble rock similar in chemical composition to Tums. Over millions of years, cracks in this karst have been widened by acidic rainwater dissolving away the limestone to make caves. The Dinaric Karst is chock-full of such caves which form a huge maze of interconnected systems deep into the earth (the limestone goes down for kilometers into the crust)!
Congeria kusceri shows its white, unpigmented flesh. Photo by Vedran Jalžić
So if you were to put all your cards into evolving to be a cave creature, the Dinaric Karst would be a great place to try it. All sorts of spooky trogloditic creatures are found down there, such as olm salamanders, cave-dwelling isopods (rolly-pollies), and more. But Congeria is of particular interest because it is a remnant of a group of mussels that was once much more prevalent across this area of Europe, once a huge mass of swamps and lakes called Paratethys. Many species of mussels in the same family as Congeria can be found as fossils in rocks covering an area reaching from Switzerland to Lake Aral in Central Asia. But over time, the climate and geography of the area changed and almost all of these species went extinct. It was once assumed that Congeria had itself gone extinct 5 million years ago, as it was only known from fossils.
Congeria kusceri. Photo by The Croatian Biospeleological Society
In the 1960s, a team exploring the caves of the Dinaric Karst were shocked to find examples of living Congeria in caves. In many ways, the story was very similar to the discovery of coelocanths in the deep sea, a fish assumed to have gone extinct millions of years ago which was found by accident. These species are remnant twigs of previously lush branches of the tree of life. Such species are sometimes called “relict” taxa, or sometimes “living fossils.” Many biologists prefer not to use that phrase, because while these creatures may closely resemble the forms of their distant ancestors, they often have changed in all sorts of ways not easily visible to the naked eye. Congeria was unknown to us, but spent millions of years evolving to improve its fitness in its cave environment. Such groups thought extinct only to reappear later in the fossil record are also sometimes called “Lazarus taxa.” Like Lazarus in the biblical story, Congeria was thought dead only to reappear much later.
It appears the species making up Congeria were trapped in Paratethys first by the rise and retreat of the Mediterranean Sea and then trapped in caves following the drying up of Paratethys. For the past several million years, they have lived only in the cave systems of the Dinaric Alps. But like Gollum, they were not preserved exactly as they previously lived in the Paratethys. While other freshwater mussels continue to frolic like merry hobbits through the streams of the Dinaric Alps above, Congeria has adapted in all sorts of interesting ways to its life as spelunking animals.
Like other bivalves, Congeria makes a living filter feeding. But in caves, they don’t have access to the photosynthetic plankton food that is the main dietary staple for typical bivalves. Congeria instead feeds on whatever detritus flows its way through the waters of the caves, and it can’t be picky. Bivalves use their gills to catch food when filtering, and Congeria has a particularly bulked up set of gills to supercharge its ability to grab these particles. But the palps, organs to select which particles to actually bring to the animal’s mouth, are tiny to the point of being almost absent. As with Gollum, who was known to gobble down the occasional goblin, Congeria can’t be selective. They have to try to eat whatever comes its way, unlike other bivalves such as my Biosphere 2 clams, which spit out many of the particles they catch as “pseudofeces.”
In evolution, if you don’t use a feature, you often lose it. Organisms can’t afford to be sentimental about organs; if it saves energy and doesn’t cost any survival ability to cut out a particular feature, it will be reduced or lost over the generations. Like other cave animals, Congeria has lost almost all pigmentation since such pigments have no purpose in the absence of light. It also has a highly reduced foot, which is used by other freshwater bivalves to move around and dig in the sediment. In the stable, low energy environment of caves, a muscular foot would be a waste of energy. It has lost its statocysts; balance stones similar in function to our inner ear used by most mollusks to sense which direction is up. Since it settles on any available surface and doesn’t move, it makes sense to strike that item from the energy budget.
Congeria living comfortably out of water, one with valves open! Source Glavaš et al. 2017
Most bivalves in the freshwater mussel Dreissenidaefamily are “r strategists“, meaning they are like guppies or mice, producing and broadcast spawning a huge number of offspring. They don’t put too much energy into each offspring, and hope a certain small percentage of them will survive each year to keep the cycle going. R strategists often don’t live very long. Such a life strategy would be tough to sustain in isolated cave systems, which are a harsh environment in many ways, particularly regarding food supply for baby Congeria. Congeria instead has evolved to be a “K strategist” like humans or elephants. It “broods” its young rather than broadcast spawning. This process, found in other bivalves from extreme environments such as polar regions, involves taking in sperm to fertilize eggs internally. The mother clam then nurtures the embryos to a certain size, first within the gills and then in special pouches in the mantle tissue. This organ is called a marsupium; these clams are like kangaroos! Brooding is much more energy-intensive than broadcast spawning, since the mother is sharing energy with her babies, but the investment ensures that each baby Congeria will reach a larger size before leaving its mother’s shell, giving each one a better chance of surviving long term. In the resource-deprived environment of a cave, a little boost to reach maturity matters! Because real estate is fairly limited in its caves, Congeria can survive in very dense aggregations with over 1,600 individuals per square meter. These mussels are surprisingly selfless and communally living for such a harsh environment.
An acetate peel of the hinge of Congeria kusceri, showing annual growth lines used to tell its age. Source: Puljas et al. 2014.
As with Gollum, Congeria has become very long-lived. Individuals have been confirmed to live almost 60 years based on growth lines in its shell, and there are suggestions that they can live much longer. In the relatively predatory-free environment of the caves, Congeria can bet on not being killed before it reaches its prime. Instead, its greatest enemy is the fickle environment of the caves, which are dependent on water flow from the surface. Their shell growth is interrupted during the summer, when the waters of its cave become much less alkaline than at other times of year, depriving them of the calcium they need to build the shell. At other times they might have to survive extended periods being emersed from the water, when levels drop during the dry season or droughts. Most bivalves can only persist for hours in such conditions, but Congeria can survive 2 months out of water! For Congeria, it makes sense to bet on being able to survive as long as it can in the spot it settled, trying to reproduce little by little over a long period of time. In this way, it is not quite like Gollum, as it has evolved to live a long time to improve its ability to reproduce and pass on its genes, rather than out of selfish worship of a ring.
Like Gollum though, Congeria is very unique and fragile. The caves in which it lives are a sensitive environment, vulnerable to overdevelopment and pollution. Manipulation of groundwater has reduced the flow of water through these caves and thus restricted Congeria‘s habitat, wiping it out from areas of some caves. Because its population is inherently restricted by the “small pond” it inhabits, it would not take a big environmental disruption to wipe Congeria off of the map. To that end, Congeria is red-listed as endangered and is protected in most of its native countries. Still, we can only hope that even now there are other populations of Congeria waiting to be discovered, hidden in unexplored caverns deep in the Dinaric Karst.





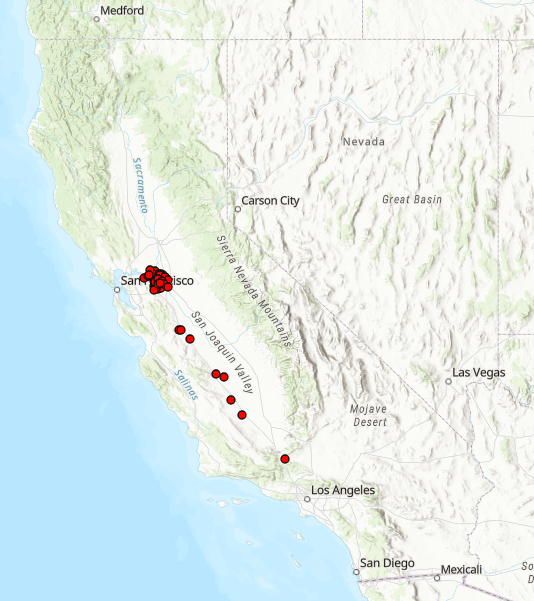






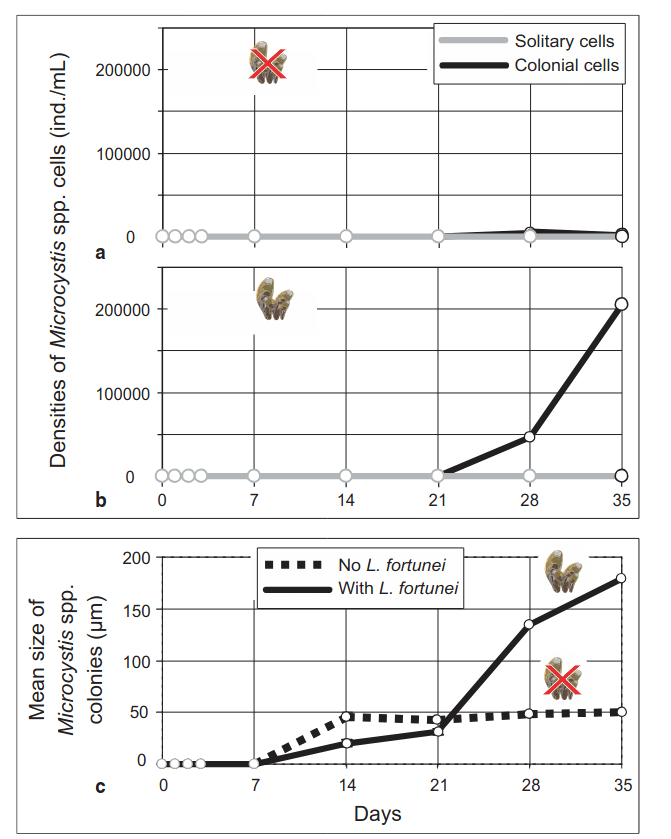

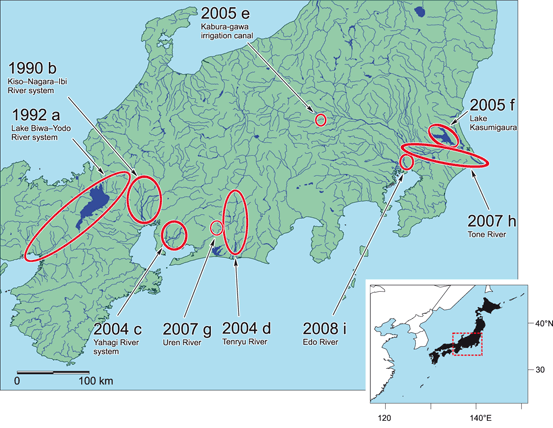
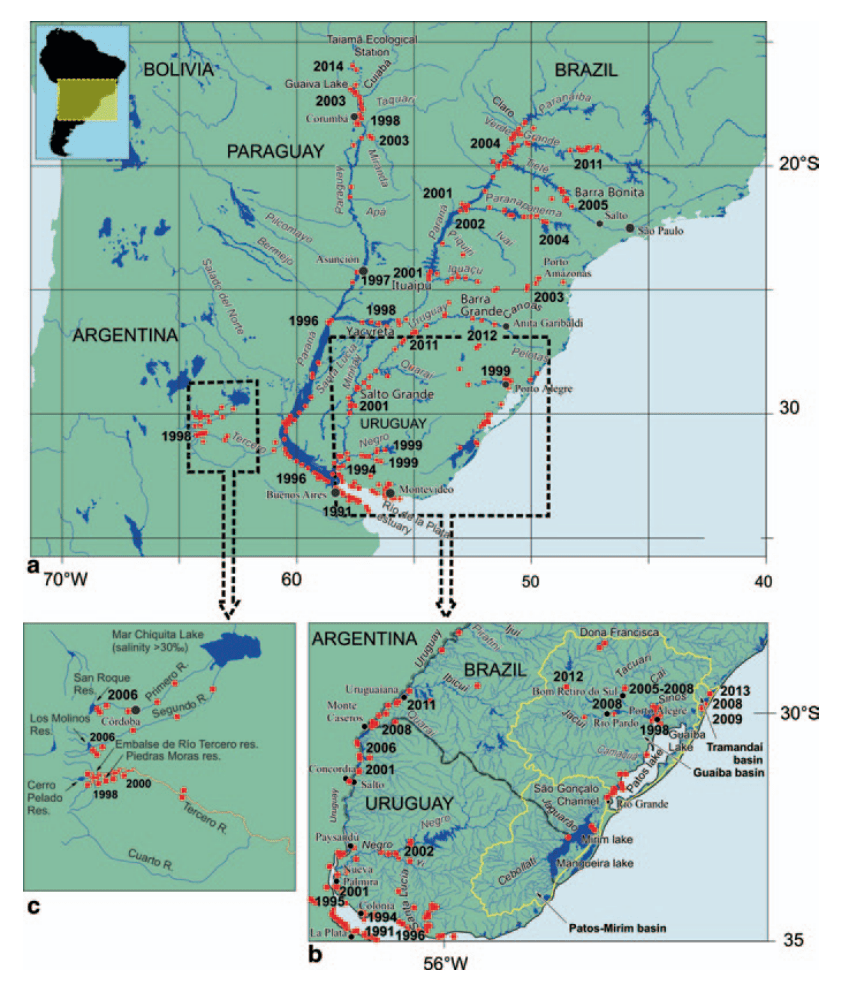
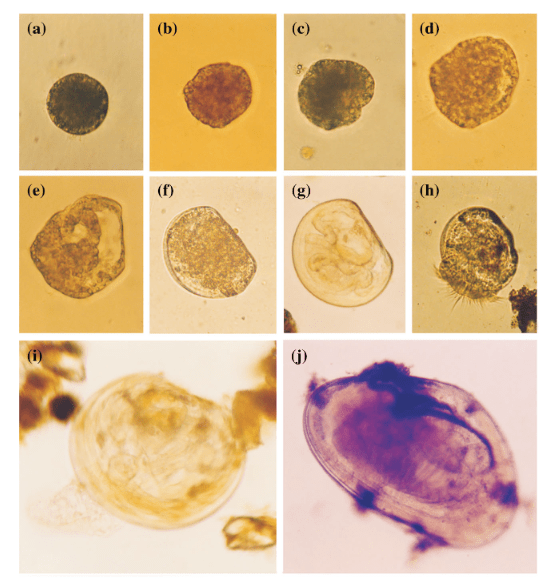



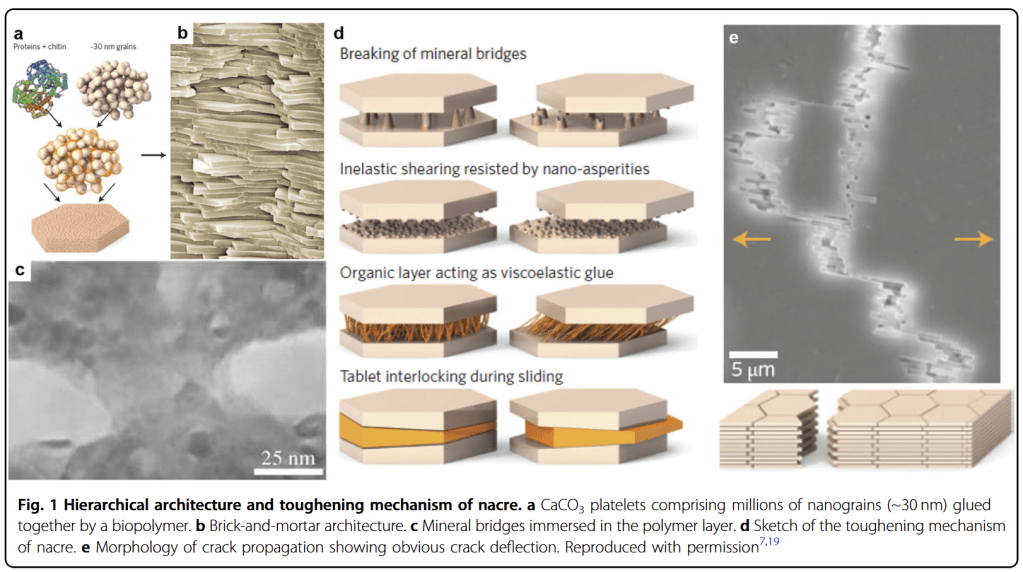


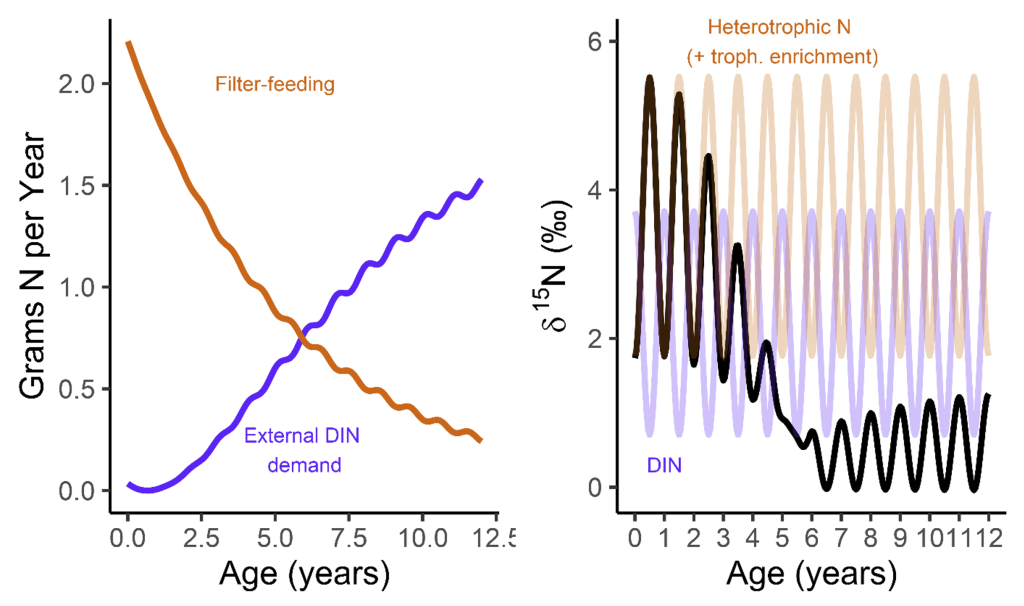




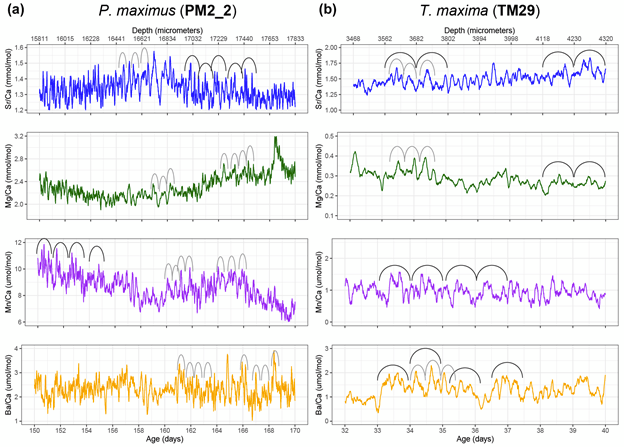
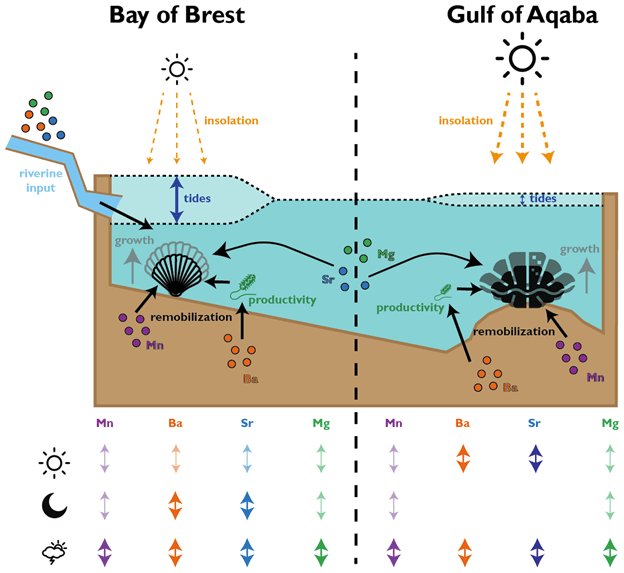



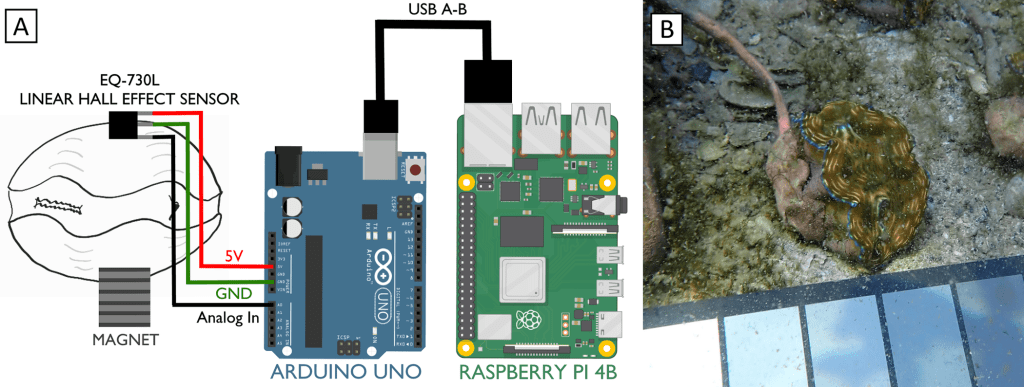


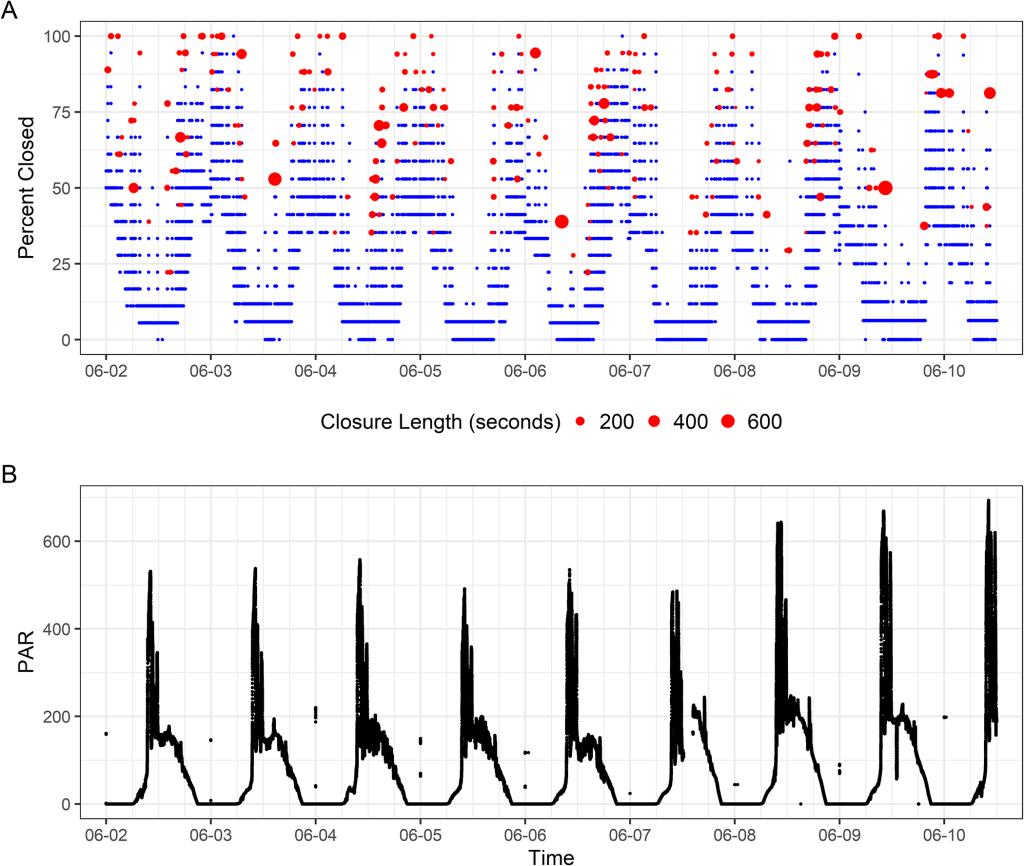


















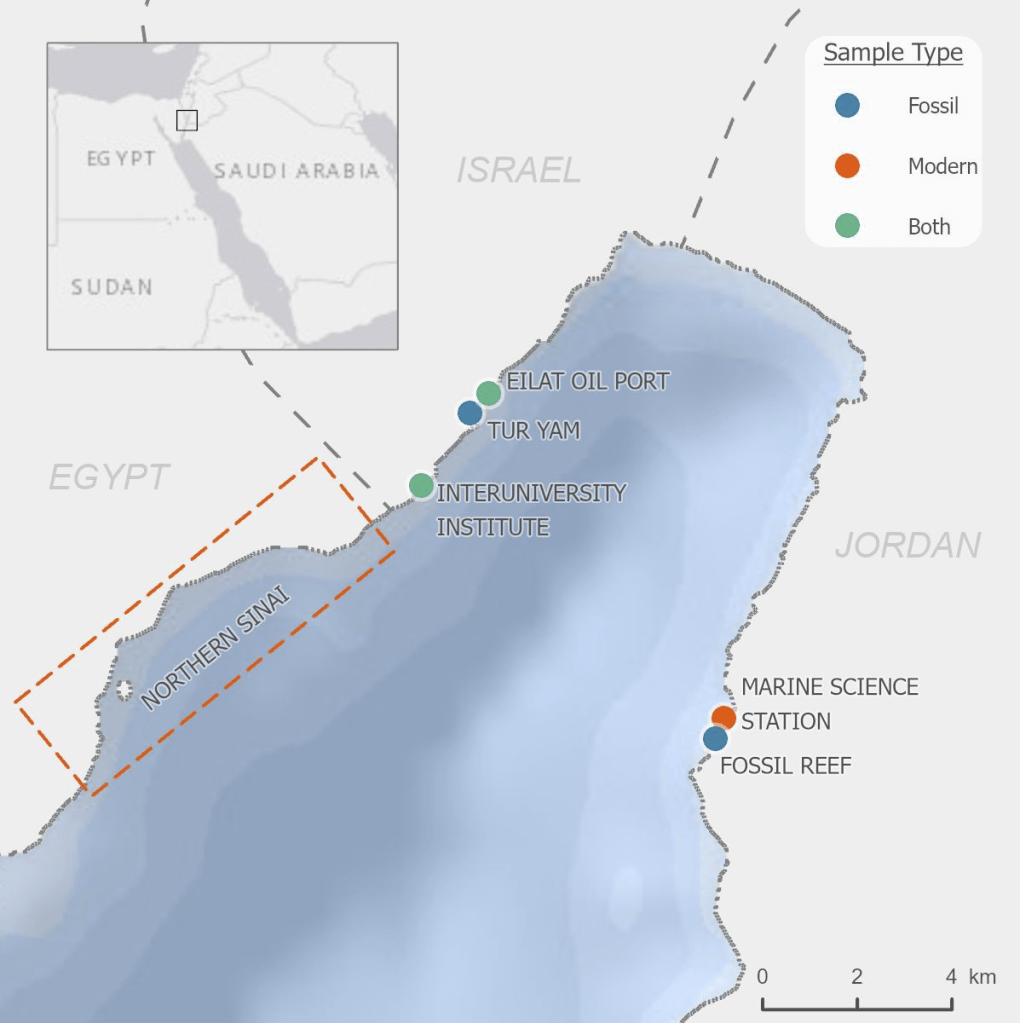

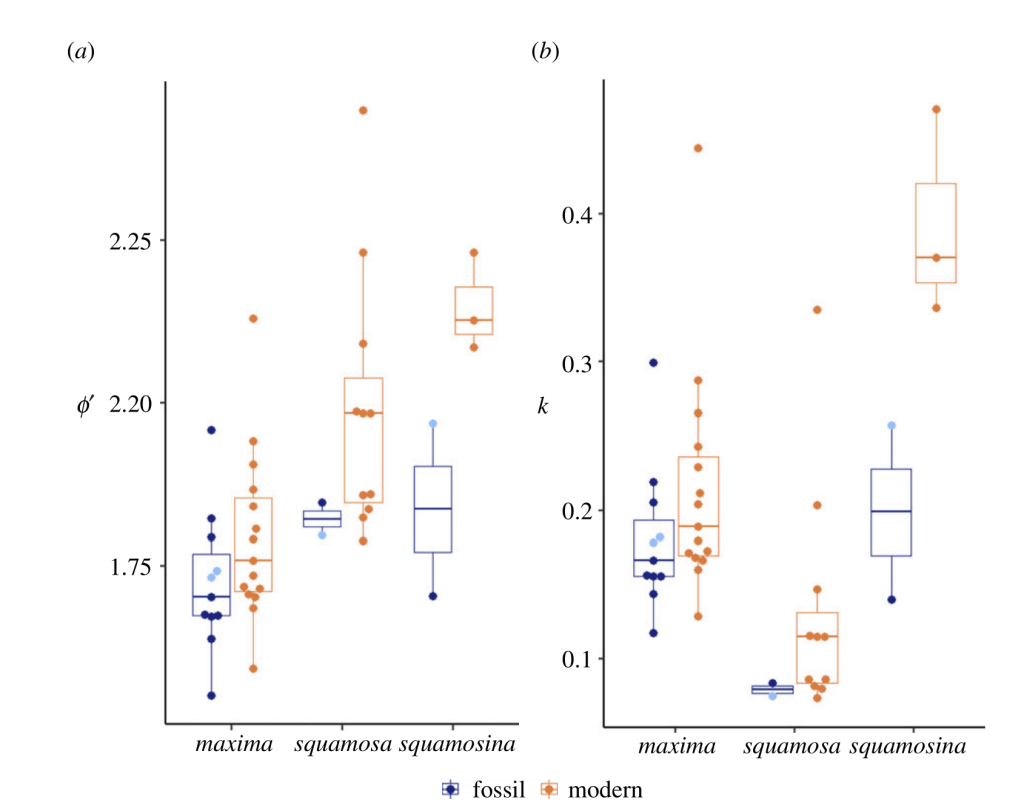
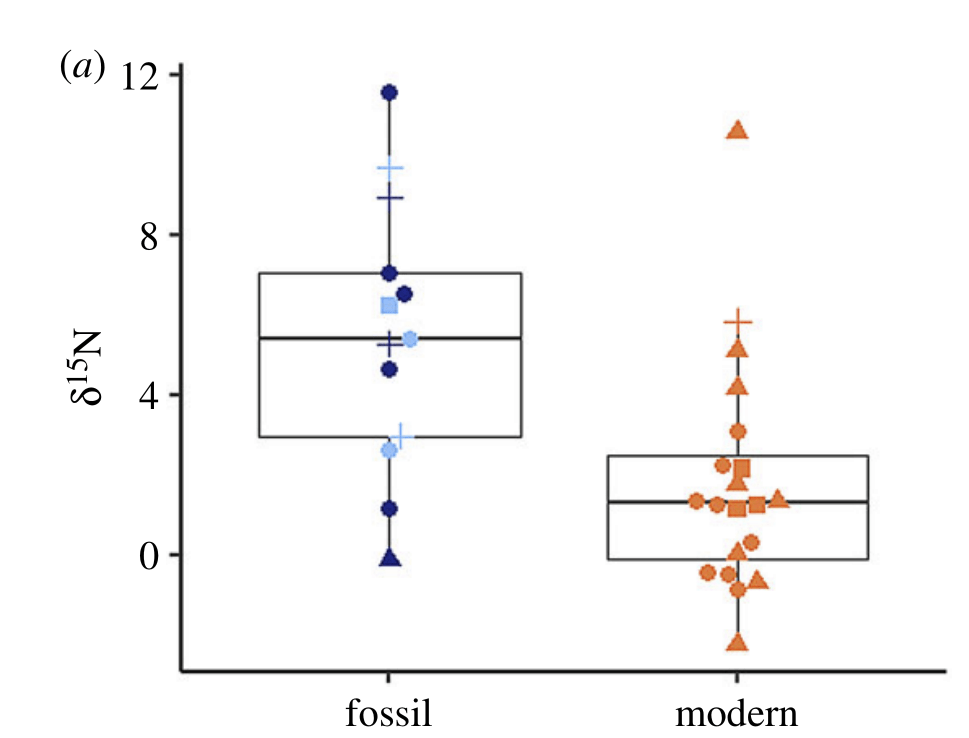









{kind=link}