

There are many types of giant clam. Not all of them are giant; the boring giant clam, Tridacna crocea, only grows to 10 cm long or so. The boring giant clam is not so named because it’s dull; its main skill is its ability to bore into the coral of its coral reef home and live with its entire shell and body embedded in the living coral. They sit there with their colorful mantle edge exposed from a thin opening in the coral, harvesting energy from sunlight like the other giant clams. When disturbed by the shadow of a human or other such predator, they retract their mantle and close their shell, encased by an additional wall of coral skeleton. It’s a clever defensive strategy, and they are some of the most numerous giant clams in many reefs in the Eastern and Southern Equatorial Pacific.
But it’s always been a mystery of how they bore away at the coral so efficiently, and how they continue to enlarge their home as they grow their shell. There are other bivalves that are efficient borers, including the pholad clams (“piddocks”) which use sharp teeth on their hinge to carve their way into solid rock, and the shipworms, which have abandoned their protective shell and instead use their two valves as teeth to burrow into wood. Both of these methods of boring are pretty straightforward.

Piddocks in next to holes that they made in solid rock. Source: Aphotomarine

Shipworm embedded in wood. Source: Michigan Science Art via Animal Diversity Web
But the boring giant clam has no such adaptation. It does not have large teeth on its hinge to carve at the coral. Such abrasion of the coral would also not explain how they widen the opening of their cubby-holes to allow their shell to grow wider. This mystery has long confounded giant clam researchers. I myself have wondered about it, and was surprised to find there was no good answer in the literature about it. But now, a team of scientists may have cracked the problem once and for all.
At the back of T. crocea‘s shell at the hinge, there is a large “byssal opening” with a fleshy foot which they can extend out of the opening to attach themselves to surfaces. Giant clams that don’t embed in coral (“epifaunal,” resting on the surface of the coral rather than “infaunal,” buried in the coral) lack this opening. The researchers suspected that the foot was the drilling instrument the clam used to create its home.

Byssal opening of T. crocea with the foot retracted. Source: NickB on Southwest Florida Marine Aquarium Society
How could a soft fleshy foot drill into the solid calcium carbonate (CaCO3) skeleton of corals? I can confirm from experience that my own foot makes for a very ineffective drilling instrument in such a setting. But T. crocea has a secret weapon: the power of acid-base chemistry. CaCO3 can be dissolved by acids. You may well have taken advantage of this chemistry to settle your acid stomach by taking a Tums, which is made of CaCO3 and reacts with the excessive hydrochloric acid in your stomach, leaving your tummy with a more neutral pH. pH is a scale used to measure acidity, with low numbers indicating very acidic solutions like lemon juice, and high pH indicating a basic solution like bleach.
Scientists are well aware of the hazards corals face from decreasing pH (increasing acidity) in the oceans. All the CO2 we are emitting, in addition to being a greenhouse gas, dissolves in the ocean as carbonic acid and gets to work reacting and dissolving away the skeletons of corals and any other “calcifying” organisms that make shells. It makes it harder for corals to form their skeletons and is already worsening die-offs of corals in some areas. The researchers suspected that the clams use this phenomena to their advantage at a small scale, lowering the pH with their foot somehow to dissolve away the coral to make their borehole.

But they needed to prove it, and that was a challenge. Giant clams can be unwilling research participants. I myself have observed this in trying to take samples of their body fluid for my own research. When they sense the presence of a predator, they immediately clam up in their protective shell. I used a small wedge to keep their shells open to allow me to take a sample of their body fluid, but the researchers working on T. crocea needed to convince the clam to place its foot on a piece of pH-sensitive foil, keep it there and do whatever acid-secreting magic allows it to burrow into coral. They would then be able to measure whether it indeed is making the water around its foot more acidic, and by how much.
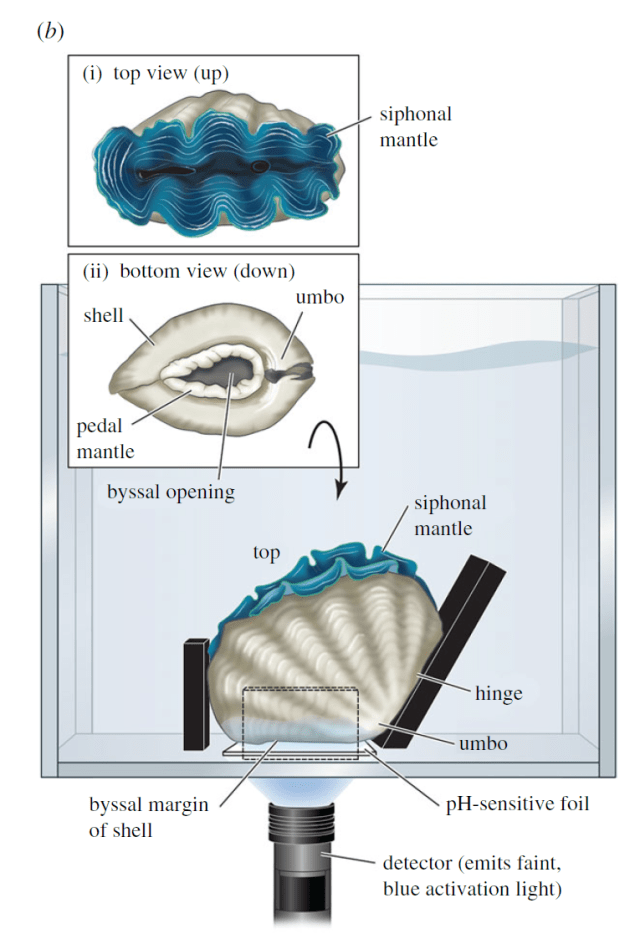
In what I can only assume was an extended process of trial and error and negotiation with a somewhat unwilling research subject, the researchers found exactly the right angle needed to convince the clam that it was safe enough to try making a coral home. But it was not in coral, instead sitting in an aquarium, on top of a special type of foil that changes color when exposed to changing pH, like a piece of high-tech litmus paper. The researchers discovered that their suspicions were correct: the clams do make the area around their feet significantly more acidic than the surrounding seawater, as much as two to four pH units lower. Where seawater is around a pH of around 8, the clams were regularly reducing pH to as low as 6 (about the level of milk) and sometimes as low as 4.6 (about the pH of acid rain). Small differences in pH can make a big difference in the power of an acid because each pH unit corresponds to 10x more protons (hydrogen ions, H+) in the water. The protons are the agent that dissolves CaCO3. Each proton can take out one molecule of coral skeleton. The clams are dissolving away coral skeleton to make holes with only their feet!
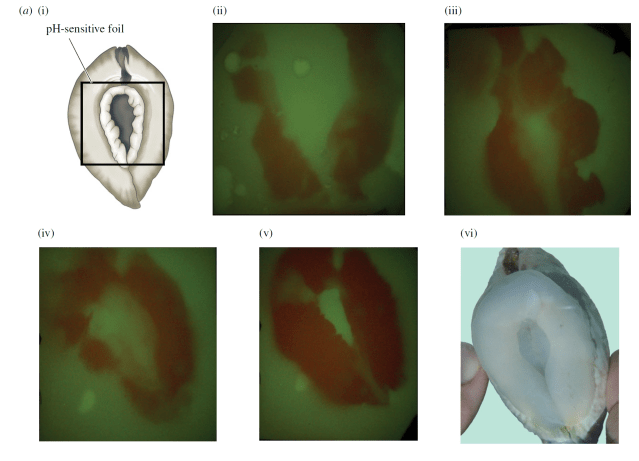
But what in T. crocea‘s foot allows them to make acid? I know that my foot does not do this, though that would be a very entertaining and obscure superpower. The researchers found the enzymes called vacuolar-type H+-ATPase (VHA) present in great quantities in the outermost cells of the clam’s feet. These enzymes are found throughout the tree of life and are proton pumps that can quickly reduce pH through active effort. Other prior researchers like the influential Sir Maurice Yonge, a legendary British marine biologist who worked extensively with giant clams, had suspected that the clams had used acid but had never been able to detect a change in pH in the seawater around the clams’ feet through more conventional methods. It was only because of new technologies like the pH paper that this research team was able to finally solve this issue. And now, I suspect other groups will want to re-investigate the importance of VHA in their study organisms. Many branches of the tree of life may be utilizing acid-base chemistry to their advantage in ways we never had previously imagined.







{kind=link}