Another year, another new paper is out, another clamsplainer to write! The fourth chapter from my PhD thesis was just published in Proceedings of the Royal Society B. This study represents five years of work, so it feels great to finally have it leave the nest. In this study, we investigated the comparative growth of fossil and modern giant clams in the Gulf of Aqaba, Northern Red Sea. Back in 2016 during my PhD, I knew I wanted to study giant clams because they are unique “hypercalcifying” bivalves that grow to huge sizes with the help of symbiotic algae living in their bodies. The clams are essentially solar-powered, and use the same type of algae that reef-building corals depend on! Unlike corals, which are the subject of a ton of research related to how they are threatened by climate change, habitat destruction and pollution, comparatively little is known of how giant clams will fare in the face of these environmental changes. Are they more resistant than corals, or more vulnerable?

I had strong reason to suspect that the clams are struggling in the face of human changes to the environment. They can bleach like corals do when exposed to warm water, and have been observed to be harmed when waters are less clear since they are so reliant on bright sunlight to make their food. But I need a way to prove whether that was the case for the Red Sea. I needed to travel to a place where fossil and modern giant clams could be found side by side, so their growth could be compared using sclerochronology. We would count growth lines in their shells to figure out how fast the grandaddy clams grew before humans were around, and compare that ancient baseline to the growth rate of the clams in the present. Giant clams make growth lines every day in their shells, giving us the page numbers in their diary so we can figure out exactly how fast they grew! We can also measure the chemistry of their shells to figure out the temperatures they experienced from the oxygen isotopes, and even what they were eating from the nitrogen isotopes.
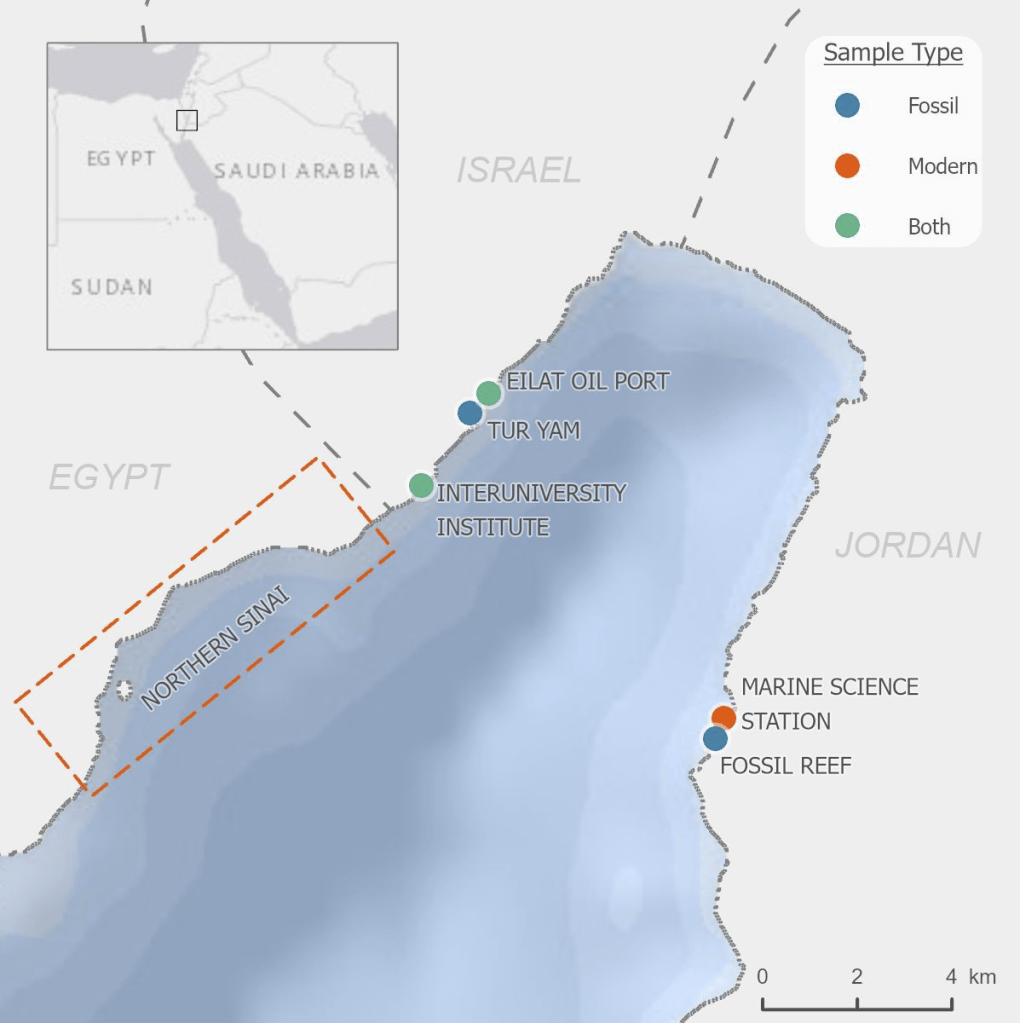
It just so happened that UCSC’s Dr. Adina Paytan was leading an NSF-funded expedition to the Red Sea in summer 2016, which represented a perfect place to do this work. There are many age dated fossil reefs uplifted onto land around the Gulf of Aqaba on the coasts of Israel and Jordan, and there are three species of giant clam living in the Red Sea today: Tridacna maxima (the small giant clam), Tridacna squamosa (the fluted giant clam), and Tridacna squamosina. Tridacna squamosina is particularly special because it is only found in the Red Sea, making it an endemic species. It is extremely rare in the modern day, with likely only dozens of individuals left, making it potentially endangered.
So I set off with Adina and two other students to live for two months in in the blazing hot desert resort town of Eilat, Israel, working at the famous Interuniversity Institute. Getting a permit from the Israeli National Parks Authority, I collected dozens of empty giant clam shells (no clams were harmed in the course of this study!) from the surf zone and from ancient reefs ranging from a few thousand years to almost 180,000 years old. I also spent a week over the border in Aqaba, Jordan where I worked with Dr. Tariq Al-Najjar, my coauthor and director of the University of Jordan Marine Science Station. Tariq is a specialist in algal productivity in the Gulf and was an excellent resource in trying to understand how water quality has changed in the area through time. He pointed out that over the years, the Gulf of Aqaba has had an increased nutrient supply far above what it received in historic times. For nearly 20 years the Gulf was subjected to excess nutrients from Israeli fish farms, which caused tremendous damage to the reefs of the area with their releases of fish waste. The farms were finally forced to close after a long lobbying campaign from Israeli and Jordanian scientists and environmentalists. But even after the farm pollution stopped, there was still increased nutrient supply from runoff and even carried into the Red Sea by dust in the form of nitrate aerosols. These aerosols are produced when our cars and power plants release nitrogen oxide gases, which react in the atmosphere to form nitrate and fall during periodic dust storms that hit the Red Sea a few times per year.
All of these sources of nitrogen are fertilizer for plankton, causing what scientists call “eutrophication.” When plankton blooms, it literally causes the water to be less transparent, which could reduce the clams’ ability to gather light and lead to them growing more slowly. At least that was my hypothesis, but I had to prove if it was true or not. So during that summer and over the next few months, I cut dozens of clam shells into cross-sections, used a special blue dye called Mutvei solution to make their growth lines visible, and took pictures of those lines with a microscope. Then I counted those lines to figure out how many micrometers the clam was growing per day.

Here I hit my first challenge: it turned out some clams were putting down one line per day as expected, but some were putting down twice as many! But the way I was using to discern between the two was to measure the oxygen isotopes of the clams’ shells, which forms a record of temperature. By counting how many lines appear between each annual peak of temperature, we confirmed some were daily and some were twice daily. But the oxygen isotope approach is expensive would not be scalable across the dozens of shells I had collected.
Then I remembered that I could measure the lines in the inner part of the clams’ shells, which are formed annually. By counting those lines and then measuring the length of the clam, I could get an approximate measure of how much it grew per year on average. This would allow me to calibrate my band-counting and discern which records represented daily lines and which were twice daily! What a relief.
So I went through all of the shells, counting lines and gathering growth info for as many shells as I could muster. It meant many hours staring at a microscope, taking pictures and stitching the pictures together, then squinting at my computer screen highlighting and measuring the distance between each growth line. I had hoped to come up with an automated way to measure it, but the lines turned out to be faint and difficult for the computer to distinguish in a numerical way. So instead I just powered through manually. When I had the raw growth data, I then transformed them to a pair of growth constants commonly used in the fisheries literature to compare growth across populations. When I put the data together across all 55 shells, I was surprised to discover that my hypothesis was totally incorrect. The clams were growing faster!
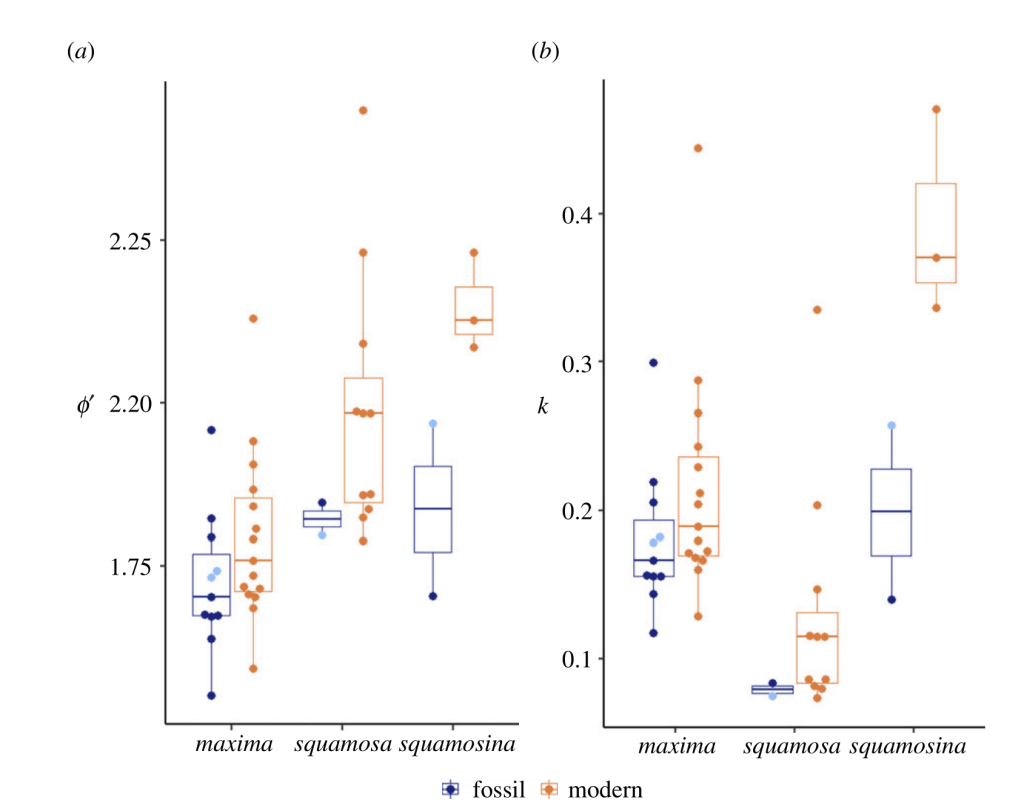
Science rarely goes according to plan. The natural world is too complex for us to follow our hunches in understanding it, which is the main reason the scientific method came about! But at a human level, it can still be shocking to realize your data says you were totally wrong. So after a few days sitting and ruminating on these results and what they meant, I remembered what Tariq and other scientists had said about nitrates. The clams are essentially part plant. They use photosynthetic symbionts to gain most of their energy. And much as nitrate pollution can fertilize plankton algae growth, maybe it could do the same for the algae within the clams! It had previously been observed that captive giant clams grew faster when “fertilized” with nitrates or ammonia. But such an effect had never before been observed in the wild. We needed a way to demonstrate whether the Red Sea clams were experiencing this.
Fortunately, the clams also keep a chemical record of what they’re eating within the organic content of their shells. Shells are a biological mineral, made of crystals of a mineral called calcium carbonate. But within and between those crystals, there’s a network of proteins the clam uses like a scaffold to build its shell. Those proteins are made of amino acids that contain nitrogen. That nitrogen comes in different “flavors” called isotopes that can tell us a lot about what an animal eats and how it lives. The ratio of heavier nitrogen-13 and lighter nitrogen-12 increases as you go up the food chain. Plants and other autotrophs have the lowest nitrogen isotope values because they use nitrate directly from the environment. For every level of the animal food chain, nitrogen isotope values increase. Herbivores are lower than carnivores. If you live on only steak, your nitrogen isotope values will be higher than a vegetarian. The same will be true for clams. If the clams were taking in more nitrogen from sources like sewage or fish farms, they would show higher nitrogen isotope values in the modern day.
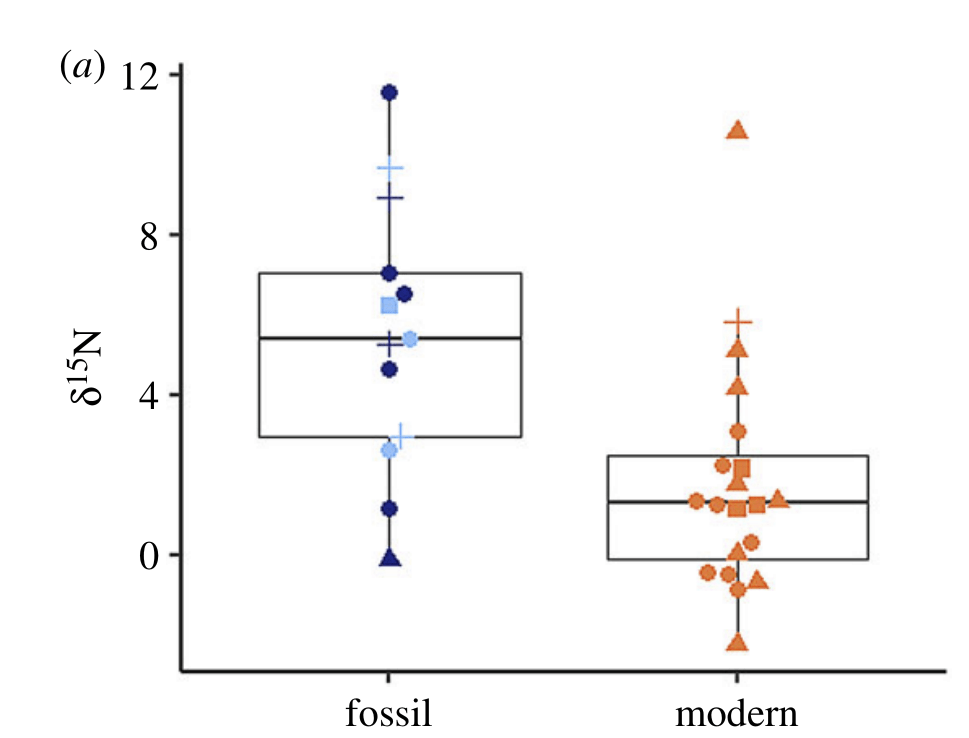
Taking bits of powder from several dozen of the shells, we worked with technician Colin Carney at the UCSC Stable Isotope Lab to measure the nitrogen isotopes of the shell material. A machine called an Elemental Analyzer literally burns the shell material to release it in a gas form. A carbon dioxide scrubber absorbs the CO2 and carbon monoxide gas, leaving only the nitrogen gas behind. That gas is measured by a mass spectrometer, which essentially separates out the different isotopes of nitrogen and tells us what fraction is nitrogen-13 or nitrogen-12. Plotting all the shell data together, I discovered that my hypothesis was…totally wrong. The nitrogen isotope values of the modern shells were lower than the fossils. The clams had moved down in the food chain, but how?

After ruling out a bunch of other explanations including the preservation of the shells, we propose that this represents a human-related change in the environment that the clams are recording. As I mentioned before, the Red Sea these days is regularly hit by huge dust storms which are conduits for nitrate aerosols. Our cars emit nitrogen-containing gases which, through a complex web of chemical reactions in the atmosphere, end up in the form of nitrate particles called aerosols. These nitrate aerosols bind to the dust delivered by strong windstorms called haboobs, which carry the dust long distances, with some of it being deposited several times of year. This deposition of nitrate has been found to form up to a third of the nitrate supply hitting the Red Sea, and was a source of nitrogen that wasn’t available to the clams in historic times. These nitrate aerosols are extremely low in nitrogen isotope value, and would be very likely to explain the lower nitrogen isotope value in our clams! If the clams ate the nitrate, their symbionts would grow more quickly, providing them with more sugars through photosynthesis and accelerating clam growth!
Some additional factors probably also have influenced giant clam growth in the region. The Red Sea historically had regular monsoon rains which likely slowed growth in fossil clams, as storms are known to do for giant clams in other areas, but such monsoons no longer reach the area. The Red Sea also had much higher seasonal range of temperatures in the past, with colder winters and warmer summers. Both factors (storms and extremes of temperature) have been previously shown to depress giant clam growth, and so the modern Red Sea may be a goldilocks environment for the clams: a consistent year-round not too cold or hot temperature.
However, as we discuss in the paper, these factors don’t necessarily mean that the clams are healthier. Faster giant clam growth has been found in other research to lead to more disordered microstructure in their shells, which would have uncertain effects on their survival against predators like fish, lobsters and humans. Additionally, a higher nutrient supply to reefs often causes the corals that build the reefs to lose out to competition from algae that block sunlight and crowd out coral colonies. If the reefs are harmed by the climatic changes that have potentially helped the clams, the clams will still lose. Giant clams are adapted to live only where coral reefs are found, and nowhere else. So more research will be needed in the Red Sea to determine if the health of clams and corals is hurt or harmed by these nitrate aerosols, and what that will mean for their long-term survival in the area.
Over the course of working on this research, the giant clams taught me a lot about life. They taught me that my hypotheses are often wrong, but that’s alright, because my hypotheses can still be wrong in a way that is interesting. I learned to go with the flow and trust the clams to tell me their story through the diaries they keep in their shells. I have followed their lessons wherever they led. Now I am doing follow-up work growing giant clams in a giant coral reef tank at Biosphere 2 in Arizona, to directly observe how the clams’ symbiosis develops and create new forms of chemical records of their symbiosis! The work described in my paper here has led to a suite of different ongoing projects. The clams have many more lessons to teach me. Thank you clams!