I am now several months into my postdoctoral fellowship at Biosphere 2 in Oracle, Arizona! I am working with Professor Diane Thompson on a project measuring the shell and body chemistry of giant clams in Biosphere 2’s huge reef tank. Our goal is to find better proxies (indirect ways of measuring) the symbiosis of these clams with the algae they farm within their bodies. The controlled, closely monitored conditions of the Biosphere 2 ocean tank represent the perfect balance between the real ocean and the more controlled environment of a lab. Using trace metals and isotopes in their shells and tissue, we can trace back the ways that clams record their own internal biology. Wild giant clams make chemical records via the growth lines in their shells, similar to tree rings. These have been the subject of many cool past studies, but there are aspects of the “language” they use to write their shell “diaries” that are poorly understood. Much like researchers used the Rosetta Stone to decode heiroglyphics, we are observing clams as they grow in order to better translate the shell diaries of their prehistoric ancestors. Doing so, we can better understand how their ancestors reacted during past periods of climate change, and identify similar bivalves in the fossil record which may have harbored symbionts.
A view of the ocean tank at Biosphere 2
I started my postdoc remotely in May. The following months were spent sheltering at home in Southern California with my mom, supervising the installation of a cohort of giant clams into the 700,000 gallon ocean tank over Zoom. It felt like a science fiction movie, watching technicians Katie Morgan and Franklin Lane from hundreds of miles away on my computer screen as they nurtured and installed the little clams in their new home. I felt like Mission Control back on earth, watching a group of space colonists work with strange alien creatures.
Some of the T. derasas in the Biosphere tank
But in August I was able to finally move to Tucson to meet these clams in person! We had three species in the first batch: Tridacna derasa, T. squamosa and T. maxima. Of the three, T. derasa (the smooth giant clam) has proven to be the most successful in the Biosphere 2 ocean tank. All of the derasa clams from May have survived and thrived, attaching themselves to the bottom with byssal threads and growing their shells, both very positive signs of clam health!
Some of our newer batch of T. derasa in the quarantine tank
So we have doubled down on T. derasa and installed 11 more individuals last week, sourced from Palauan clam farms via a reef supply company in Florida called ORA. They are currently in a shallow quarantine tank where we will monitor them for disease and unwanted hitchhikers before introducing them to the broader Biosphere tank.
The workers at Biosphere 2 are very creative problem solvers. Giant clams need intense amounts of light to sustain their symbiotic algae and create food for themselves, a quantity of light higher than is available in the current Biosphere tank. To provide a light supplement, the engineering team at Biosphere 2 constructed a floating lighting rig with hanging LED lighting, right over the lagoon where we have the clams!
The lighting rig glows with a blue light as the sun goes down outside the Biosphere
To make sure the clams have enough light, we installed a Li-Cor light sensor to measure the exact amount of photons (light particles) hitting the clams over the course of a day. The light is measured in units of micromoles of photons per meters squared per second. A mole is 6.02 * 1023 particles, and other clam experts like James Fatheree have suggested that the clams need light levels of at least 200 micromoles/m2s to make enough food for themselves. That’s 120,400,000,000,000,000,000,000 light particles we need to hit every square meter of their habitat every second. The clam channels as many of those photons as it can to its algae residing within tubes in its tissue. The symbionts use it in photosynthesis to make sugars, which they share with their host. A well lit giant clam is a happy, well-fed giant clam! But because the glass dome of Biosphere eats up some of the light, and plankton and floating particles in the seawater eat up another portion, we use the lights to make sure the clams have the boost they need to maintain their symbiosis like they would in the clear, shallow waters of a tropical coral reef.
The Li-Cor sensor floats above the clams, telling us how much light they’re getting
Much like a new dad might read parenting books to get ideas for baby care, I am always poring through the literature trying to figure out how to maximize the growth of these clams. Dr. Fatheree is kind of like Dr. Lipschitz from Rugrats, except unlike the suspect childcare advice in the show, this real-life giant clam advice is very valuable. Like human babies, these clams can be a challenge! The clams sometimes decide to move around and get themselves into trouble, requiring us to rescue them if they get trapped behind a rock or under a pile of sand. So I have had to do a fair amount of clam-herding during my time here.
We are growing the clams for science, and there will be data to collect. We will be monitoring data like the trace metal chemistry of the clams’ tissue and shells, the color of their mantles, and the pH, temperature and oxygen levels of their environment, all to relate together to make the best clam record of their environment possible. So far, I have been snorkeling in the tank every couple days maintaining their setup. Next week, I will dive in the Biosphere tank for the first time to collect data on their shell chemistry! I have other projects in the works to measure their valves opening and closing using magnetic sensors, and to measure their color changes through time through computational photography.
That brings me to what I’ve found to be the coolest part about Biosphere 2: the people. Something about this place attracts creative, brilliant, can-do people who solve problems on the fly and are always jumping into the next project. It has been a privilege to learn and pick up technical skills from them in the brief time I’ve been here. This place is really like a space colony out of The Expanse or Silent Running. There are endless valves, pipes, tanks, exchangers and other hardware needed to keep Biosphere 2 running. Getting to witness the technical competence behind the whimsical solutions the staff comes up with, like the floating light rig, has been the most exciting part of this job for me. Everyone has a deeply ingrained curiosity and passion for science that is inspiring to see; they are as interested in my clams as I am in their corals, tropical plants, and geochemical experiments. I would argue that the human team behind Biosphere 2 is a bigger treasure than the unique metal-and-glass structure they work under, and I look forward to seeing the results all of the collaborations we have in the works!
Another one of my PhD chapters is published in the journal G-cubed, resulting from work I did in the summer of 2016 in Israel and Jordan around the Red Sea. This is my first geochemistry article in a journal, so it is a big deal to me! I thought I’d write up a clamsplainer about what I was looking for and how we went about achieving the paper.
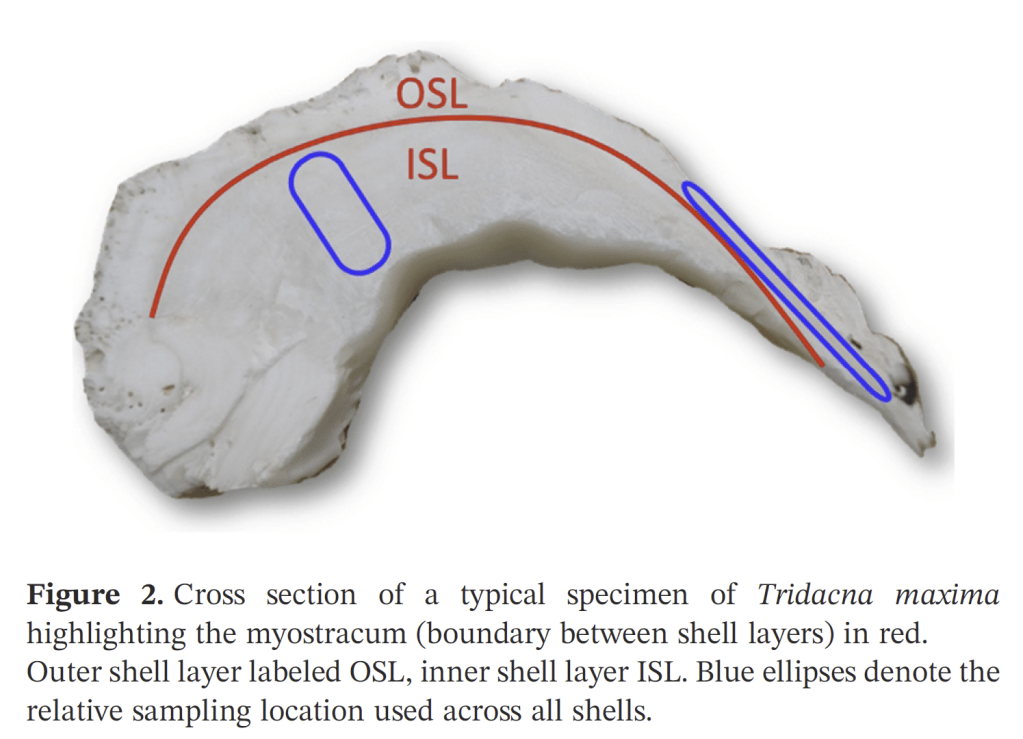
A slice of giant clam shell. You can see the difference between the inner and outer layers. The inner layer has visible annual growth lines.
I study the chemistry of giant clam shells. You might already be familiar with the concept of tree rings, a field called dendrochronology. It’s like reading the diary of a tree, where every “ring” is a page in the record of its life. The related field of sclerochronology looks at rings in the hard parts of shelled organisms. We can count those rings to figure out the ages of clams, or their health, and we can measure the chemistry of those rings to understand the temperature the clam grew at, and even what it ate.
A giant clam growing on the reef flat in Eilat
Giant clams are bivalves of unusually large size which achieve a very rapid growth rate through the help of symbiotic algae in their flesh. The clams are farmers, and their crop is inside their tissue! They grow their shells very quickly (sometimes up to 5 cm a year, equivalent to if a six foot tall man grew a foot every year from birth), and live a very long time, up to 100 years (their growth slows later in life). A whole bunch of talented researchers have measured the chemistry of giant clams all around the world to reconstruct past climate and even measure historic storms!
If we want to understand the ecology of a forest, we can’t measure just one tree!
But if you come back to the analogy of tree rings, we essentially have measured the rings and chemistry of individual “trees” in a bunch of different places, but don’t have as good an idea of how the chemistry varies within a “forest” of giant clams in a particular place. In our new study, we set out to describe exactly that, focusing on the Northern Red Sea.
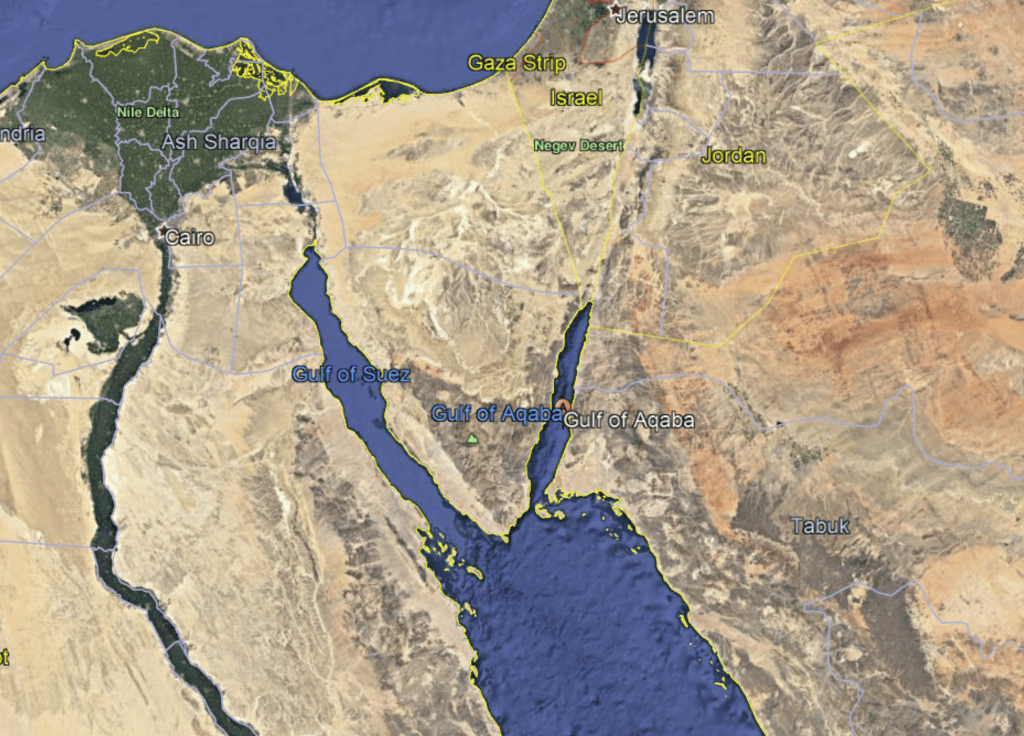
A map of the Northern Red Sea. The right “toe” is the Gulf of Aqaba
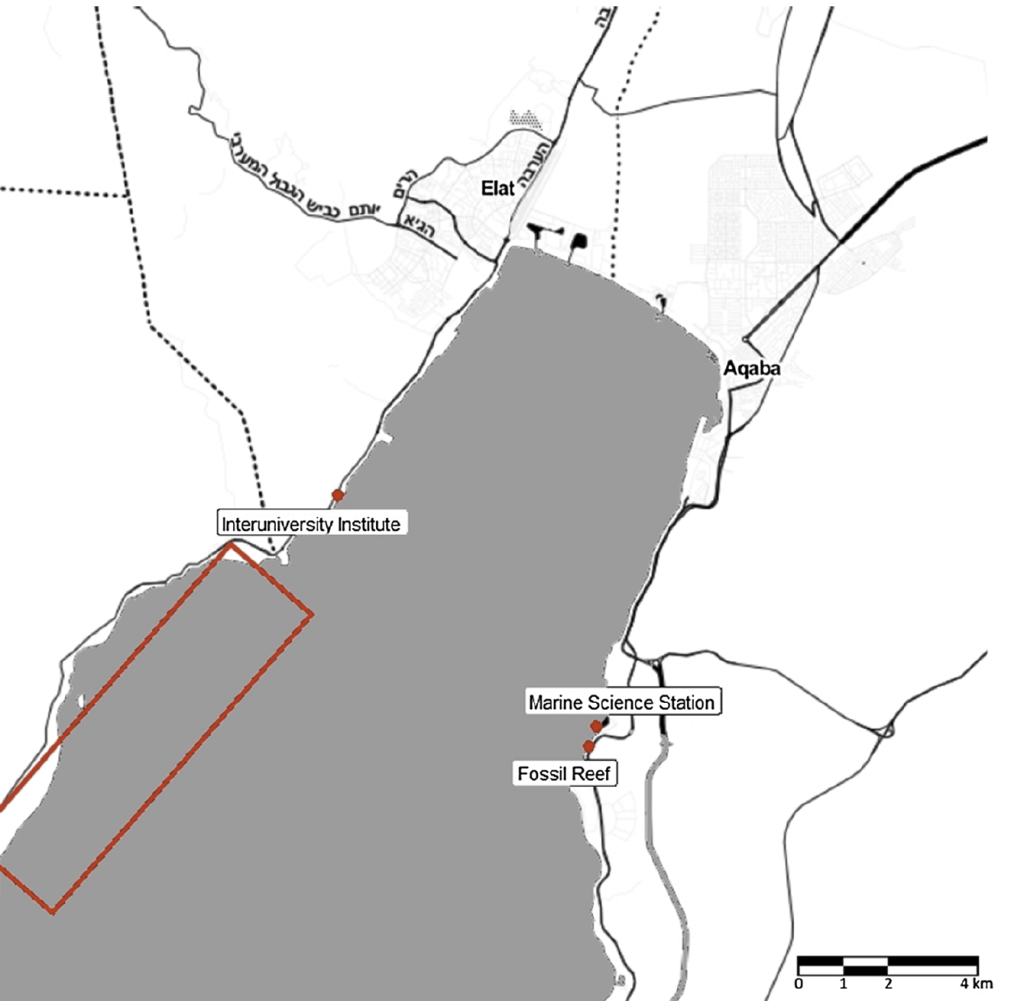
Sites where we sampled shells along the northernmost tip of the Gulf of Aqaba
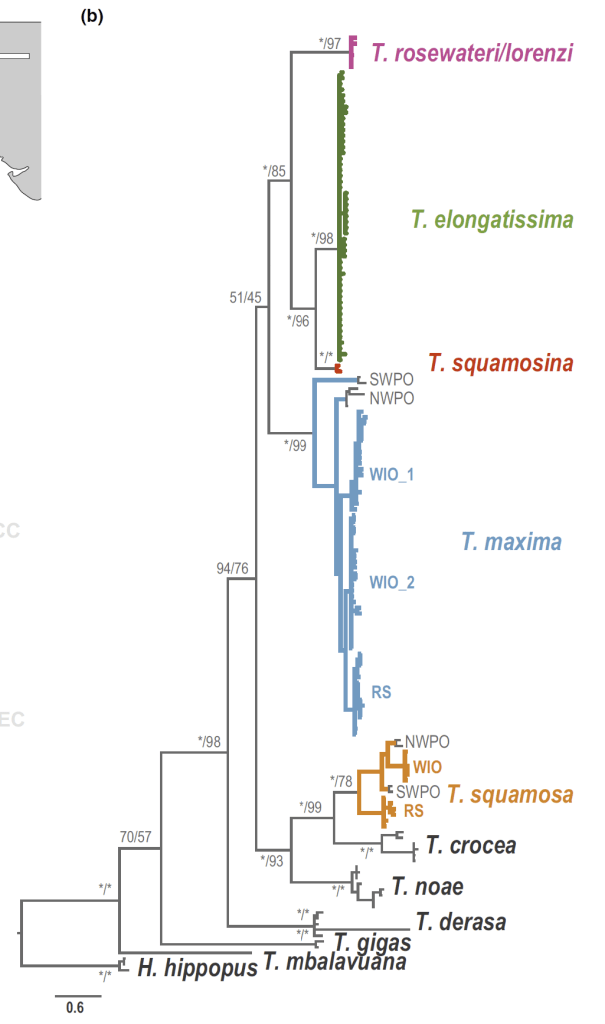
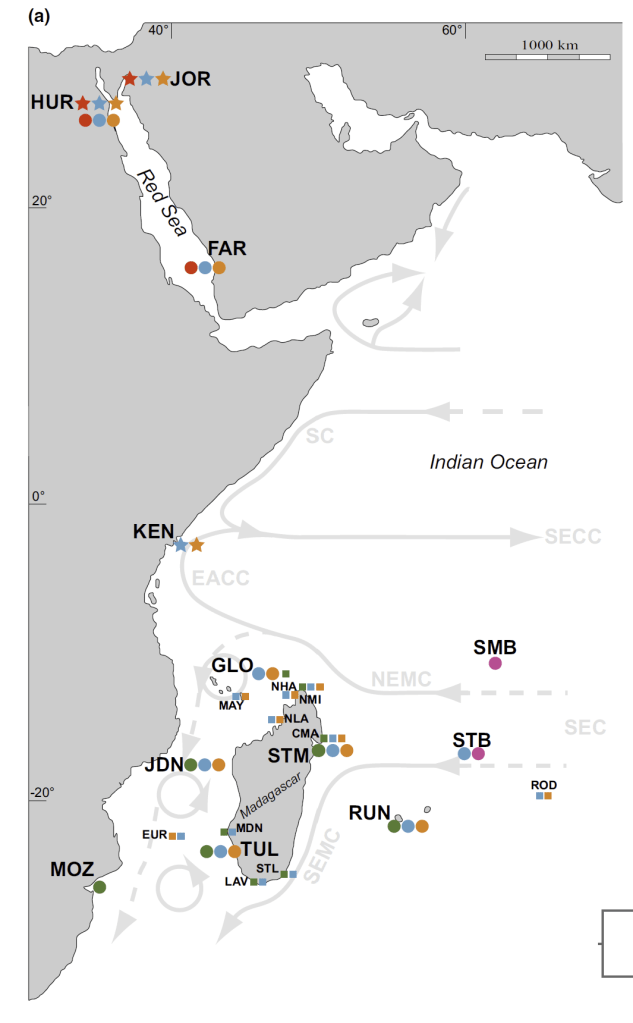
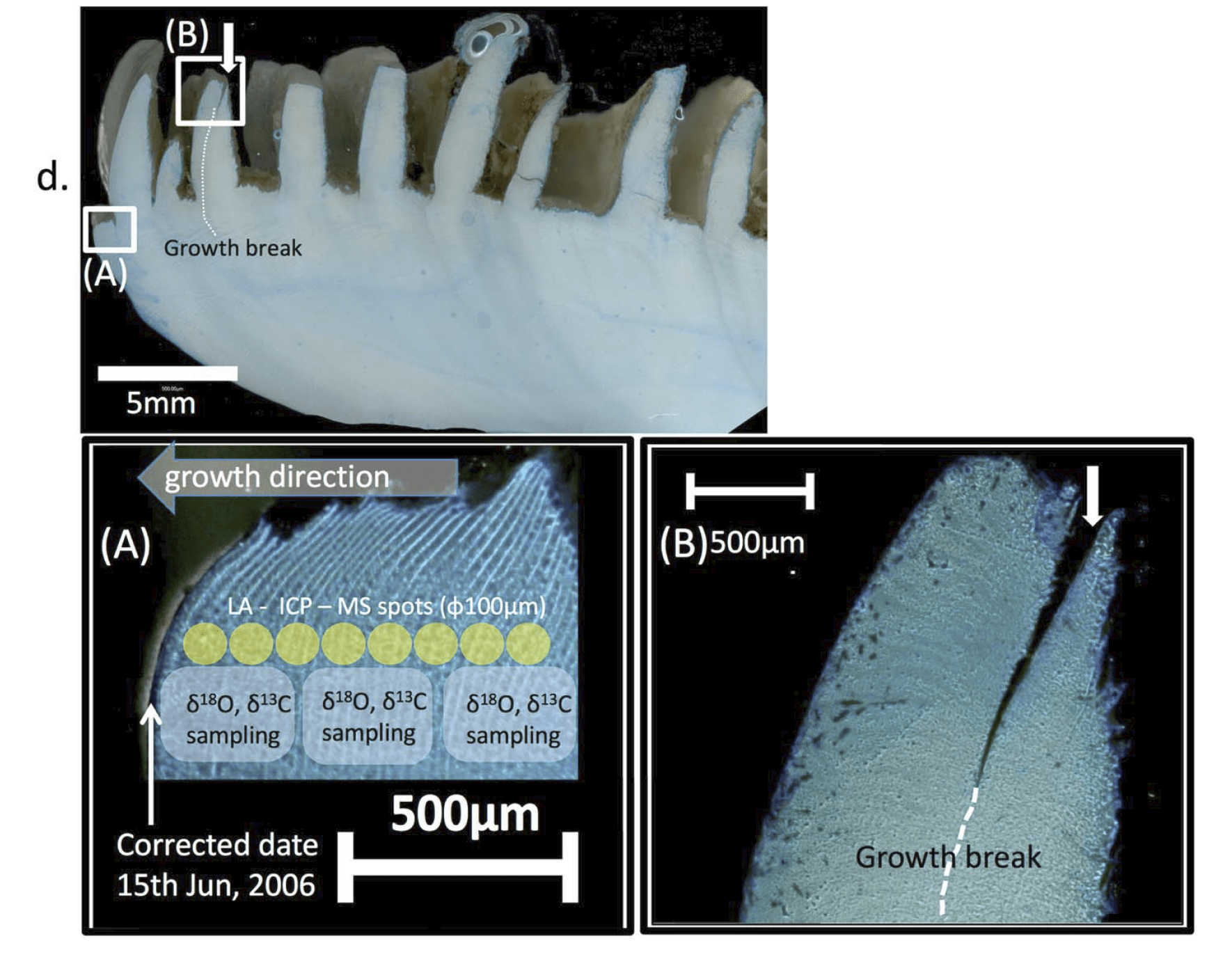
The Gulf of Aqaba represents the northernmost toe of the Red Sea, bordered by Egypt, Israel, Jordan and Saudi Arabia. It hosts some of the northernmost coral reefs in the world, aided by tropical temperatures and clear waters due to the lack of rainfall in the surrounding deserts. Here, we can find three species of giant clams including the small giant clam Tridacna maxima, the fluted giant clam T. squamosa and the very rare T. squamosina, which is found only in the Red Sea and nowhere else (as far as we know). In summer 2016, I went all around the Gulf of Aqaba collecting shells of clams from the beaches, fossil deposits, and even were able to work with shells confiscated from smugglers at the Israel-Egypt border. We cut these shells into slices and used tiny drill bits to sample powder from the cross section of their shells, which we could then conduct geochemistry with! We sampled large areas in bulk from the inner and outer portions of the shell (more on why later) using a Dremel tool, and also sampled more finely in sequential rows with a tiny dental drill bit (same brand your dentist uses!) to see how the measured temperatures varied through seasons. By “we”, I mean my coauthor and friend Ryan Thomas, who spent every Friday morning for several weeks milling out most of the powder we needed for this study. This data became part of his senior thesis at UCSC!
Two giant clams thriving on the shallow reef near Eilat, Israel
What kind of chemistry did we measure? The shells of clams are made of calcium carbonate, the same stuff Tums is made of. Calcium carbonate contains one calcium atom, one carbon atom, and three oxygen atoms. It turns out that all of those atoms come in “flavors” that we call isotopes, relating to the weight of those atoms. When you take the shell powder and put it into a machine called a mass spectrometer, you can figure out the proportions of isotopes of different elements present in the samples
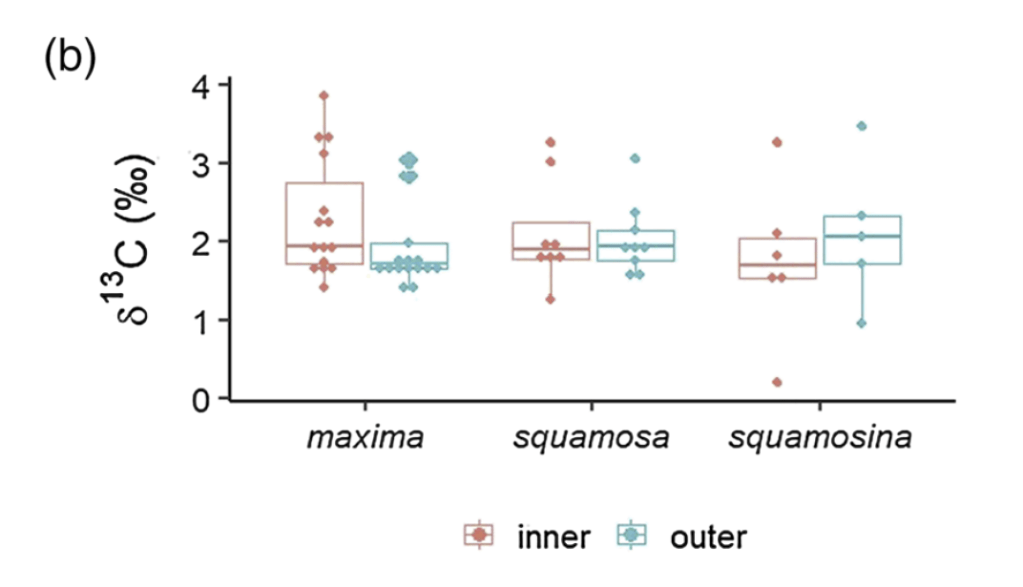
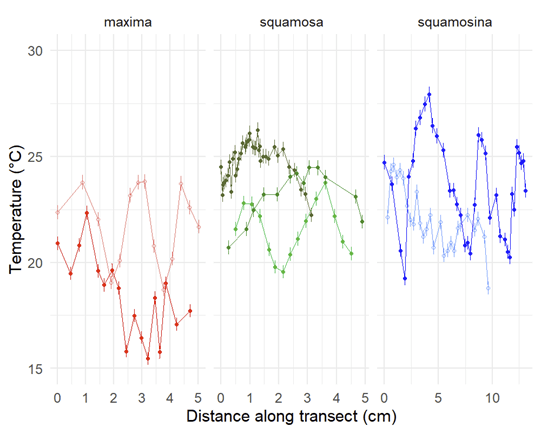
The first isotope “flavors” we were interested were carbon-12 and carbon-13. The ratio of the two is thought to relate back mostly to the action of the algae inside (its symbionts) and outside the clam’s body (the floating algae the clam filters out of the water as an additional meal). This happens because as algae take carbon from the environment and bind it into sugars through photosynthesis, they naturally weight the dice in favor of carbon-12 making it into the sugars. So carbon-13 is left out in the water, and potentially in the clam’s shell. When photosynthesis is more active, it would leave the shell with proportionally more carbon-13. At least that’s what other researchers have confirmed happens in corals, and suspect happens in clams. In the world of isotope chemistry, this phenomenon is called “fractionation,” when a process causes isotopes to form fractions separated by mass. We wanted to test if that was true for giant clams, and could do so by comparing T. squamosina and T. maxima, which have more active photosynthesis, to the less photosynthetic T. squamosa.
Comparing carbon isotopes across different species and shell layers. The results are fairly flat all the way across.
It turns out that the more symbiotic species don’t have more carbon-13 in their shells. We set out several reasons that might be the case, including that the symbionts of these clams are actually more carbon-limited than many researchers might expect. Essentially, the algae lack an excess of carbon atoms to choose from, so they can’t be picky with which isotopes they use to make sugars. Therefore, the fractionation effect weakens and becomes possibly too subtle to manifest in the shell, even in the best-case scenario of three closely related species living the same area. This represents what I’d term a “null result.” We had a hypothesis and we demonstrated that hypothesis was not the case in our data. It was important to publish this result, because other researchers will know not to try the same thing. This means that when we try to search for evidence of symbiotic algae in fossil clams, we will likely need to use other types of chemistry to figure it out. But don’t worry, as finding such a “smoking gun” for algal symbiosis in fossil bivalves is part of my life’s work! I have a few projects in the works looking for exactly that kind of evidence! 😉
A look at how temperatures measured via oxygen isotopes vary through the lives of the animals. This is how scientists can use very old shells to figure out how temperatures varied through a year in prehistoric times!
But we had additional data we collected in addition to the carbon isotopes which actually turned out to provide some interesting results. This other type of measurement regarded the oxygen isotope ratio of the shells. Previous research has shown that the ratio of oxygen-18 to oxygen-16 in carbonate skeletons directly relates to temperature, a principle that has birthed a field known as paleothermometry. There are thousands of papers which use shells of corals, clams, cephalopods, microbes and more to reconstruct temperatures in ancient times. Giant clams have proven to be effective weather stations going all the way back to the Miocene epoch, millions of years ago! Because they grow so quickly (putting down a new layer every day), live for a long time, and don’t stop growing, they form very complete, high-resolution, and long records of past climate.
But no past studies had ever compared different species of giant clams from the same place. There would be interesting new lessons to draw from such a comparison, including seeing if one species preferred to grow at warmer parts of the reef. As complex, three dimensional structures, there are many remarkably different micro-environments throughout a reef, from the hot, sun-exposed reef flat and crest to the cooler, current-swept, deeper fore-reef. Do any of the species of giant clams show a consistently higher temperature than the others, and what would that mean if they did?
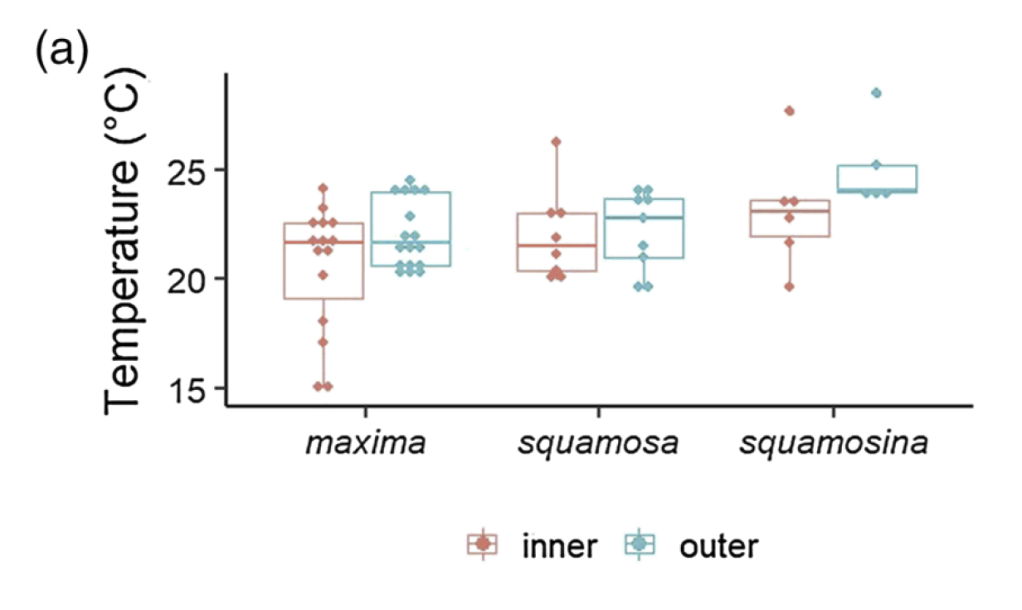
T. squamosina records higher temperatures than the other species. Outer shell layers also record higher temperatures than inner shell layers. More on that later in the post 😀
It turns out that the rare T. squamosina, only found in the Red Sea, does record a higher average temperature, almost 3 degrees C higher than the other two species. This is of interest because this species had been proposed by prior researchers to only live on the sun-drenched reef crest, at the shallowest part of the reef. We believe these results corroborate that observation. The previous research on the habitat of T. squamosina was limited to a single study which only was able to find 13 live animals along the coast of the Red Sea. But by independently confirming this life habit, we can ask further questions that may be borne out by further research.
An example of T. squamosina showing signs of possible bleaching (light parts at the center of its body).
Being restricted to the shallowest waters, is T. squamosina at greater risk of harvesting by humans along the shores of the region than its counterparts? Illegal poaching of giant clams along the Red Sea is believed to be a major stressor on their population size in the area. Could this explain why T. squamosina is so rare today, despite being proposed to have been more common in the past? In addition, being restricted to the top few feet of depth in the water could leave the species more vulnerable than the others to atmospheric warming. As with corals, when giant clams overheat they will “bleach”, expelling their symbiotic algae as a stress response. While the clams can recover, it is sometimes a fatal form of stress that leads to their death.
An excellent cartoon of the different shell layers in giant clams. From a peer of mine who also studies them, Michelle Gannon!
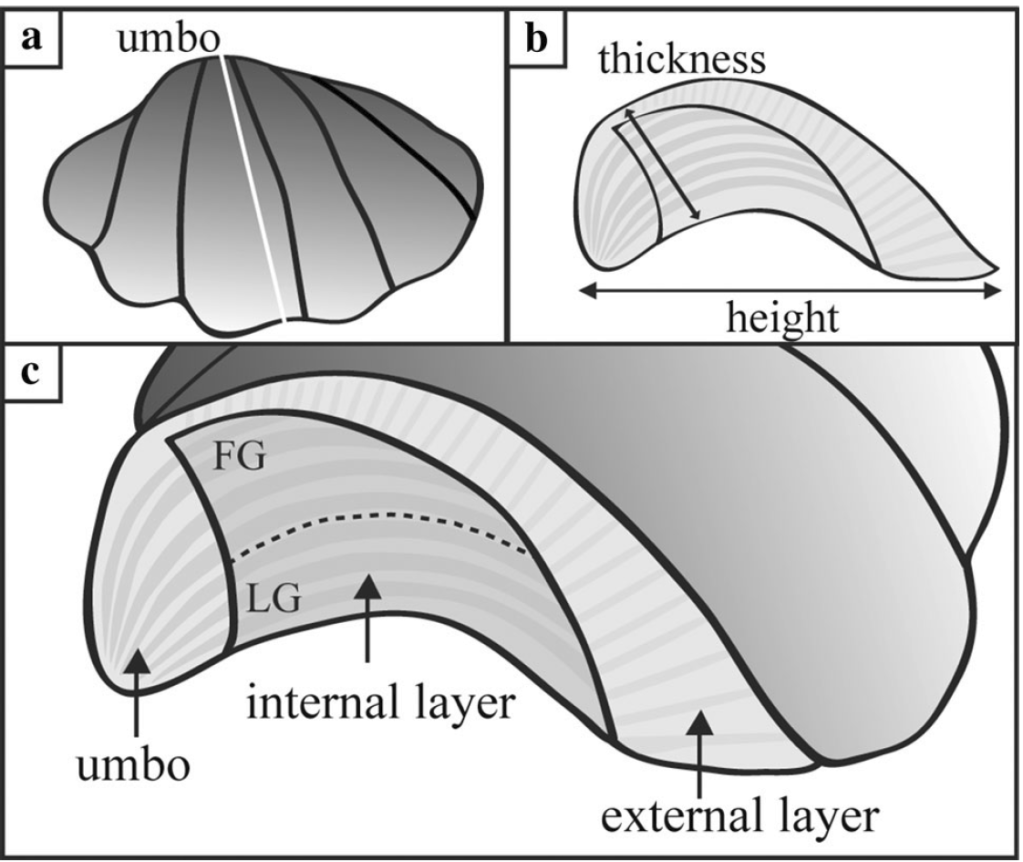
More research is needed to answer those questions. But the last aspect of this study relates to what is happening inside of the bodies and shells of the clams themselves. Giant clam shells have two layers. The outer layer grows forward away from the hinge, increasing clam’s length. The clam also makes an internal layer, growing inward to thicken the shell and add weight. We can read the growth lines of the clam’s diary within either layer, and different studies have used one or the other to make records of climate change. But very few studies have compared the two layers of the same individual. Do they record the same temperatures? Figuring it out would be important to determine how studies with just the inner layer or outer layer can be compared to each other across time and space.
A vividly blue example of the small giant clam, T. maxima. From user arthur_chapman on iNaturalist
In our studied clams, it turns out that the outer layer records warmer temperatures on average than the inner one! After ruling out other possible explanations behind this difference (the details are complicated and hard for even shell nerds to wrap our heads around), we settled on the idea that the outside of the clam is indeed warmer on average than the inside! This means that the outer layer, recording temperatures of the outer mantle, is indeed forming at a higher temperature than inside! Why is this?
Unlike us, clams are ectothermic. They generally stay the same temperature as their surrounding environment and don’t use their metabolism to generate internal heat. But that doesn’t mean that the clam doesn’t have hotter and cooler spots in its body. It makes sense that it would be hotter at the outer part of its body, facing the sun, as the solar rays hitting its outer mantle would then radiate out again as heat. The outer mantle is also darker in color than the inner mantle, allowing it to absorb more solar energy, much as you might feel hotter wearing a darker t-shirt in the sun than a white one. Photosynthesis itself produces a warming effect, a phenomenon known as non-photochemical quenching, and so the outer mantle, which contains the vast majority of the symbiotic algae, may be partially warmed by the activity of the symbionts!
More research is needed to confirm if this is true. As of yet, no researcher has ever stuck a temperature probe in multiple parts of a clam to see if the outside of it is indeed warmer than the inside. But until that day, it is interesting to think of how this would influence comparisons of diaries from the inner and outer layers of different bivalves. The effect is on the small side, so it doesn’t really mean one layer or the other should be preferred for future shell-based studies of climate change. But it could be an additional aspect to consider in the future as a way to record temperature differences within the body of an animal, and look into how those differences influence its overall level of stress.
Examples of juvenile smooth giant clams, T. derasa, that we’re growing at Biosphere 2. Photo by Katie Morgan.
So I hope this long explanation of my paper helps you to have a better idea of the work I did during my PhD thesis. There were other aspects to the paper that are too wonkish to get into here, particularly concerning the correlation we found between carbon and oxygen isotope ratios, but if you have questions or want a copy of the PDF, please message me! I have more clam papers in the pipeline, and my new postdoc at Biosphere 2 involves growing three species of giant clams in a controlled environment, where I hope to answer some of the physiological questions I mentioned above! But until then, stay hinged and happy as a clam (as much is possible in this chaotic time), and take comfort knowing there are colorful bivalves out there all at this very moment, harvesting sunlight for food and growing huge shells.
I have always been fascinated by scientific discoveries that are hanging right in front of our noses. Cryptic species are one such surprise. Sometimes, researchers using genetic sequencing are surprised to discover that a group of animals that all look the same from the outside are actually reproductively isolated from each other; separate twigs on the tree of life. This surprise has happened over and over in the history of natural science.
It turns out such puzzles are frequent among the giant clams. These unusual bivalves are specialists in coral reef environments, growing to large size with the help of symbiotic algae that create sugars through photosynthesis. Within the genus Tridacna there are ~10 accepted species which vary in size, shape, color and mode of life.
Tridacna squamosina (right) sitting next to the small giant clam T. maxima (left) on the Israeli Red Sea coast
I specialize in the three species known (so far) from the Red Sea, including the small giant clam Tridacna maxima and the fluted giant clam T. squamosa, which are both found worldwide, all the way from the Red Sea to down past the equator along the Great Barrier Reef. The third local species, T. squamosina is more unusual, so far being only known from the Red Sea (an endemic species). T. squamosina is an example of a cryptic species, having previously been assumed to be a local variant of T. squamosa. It looks pretty similar, with long scutes (flap-like appendages) protruding from its shell, thought to help stabilize it on the flat bottom of loose coral rubble. But unlike T. squamosa, T. squamosina lives exclusively at the top of the reef in the shallowest waters closest to the sun. It has a very angular, zig-zag pattern in its plications (the wavy shapes at the edge of the shell) and a characteristic pair of green stripes where the soft tissue meets the edges of the shell. The soft tissue is covered with warty protuberances.
Pictures of details of T. squamosina from Richter et al. 2008
It was only first described in detail in the early 2000s, when an international team of researchers figured out using genetic sequencing that it was a distinct species and named it T. costata. They noted that in their surveys all around the shores of the Red Sea, they only found 13 live specimens, making it an extremely rare and possibly endangered species. Fossil specimens on local reefs appeared to be much more common, suggesting it had a much larger population in the past. Then in 2011, another team at the Natural History Museum in Vienna discovered a shell of one had been forgotten in its collection for over 100 years. Rudolf Sturany, the researcher on the 1895 research cruise who had originally collected the clam, had called it T. squamosina.
The T. squamosina shell in the collection of the Museum of Natural History in Vienna (from Huber and Eschner, 2011)
In taxonomy (the science of naming and classifying organisms), the first team to name the species wins, so the name T. costata was synonymized (retired) in favor of the earlier name T. squamosina, which became the name of record. It must be annoying to spend so much time working to name a species and then discover you had been scooped over a century before! But such is science.
A mystery clam thought to be T. squamosina, later identified as T. elongatissima found off of Mozambique by iNaturalist user bewambay
The strange part was that there were some murmurs over the last few years that T. squamosina was not only found in the Red Sea, but also had been seen along the coast of Africa as far south as Kenya, Mozambique and Madagascar. Divers and snorkelers had taken pictures of a giant clam that did indeed look strangely like T. squamosina, with a zigzag shell opening and green stripes at the edge of its tissue. But some aspects of these individuals seemed off. In the Red Sea, T. squamosina lives freely, not embedded in the coral as these pictures showed, and the geometry of the angles of the shell seemed a bit different. It also would be difficult for T. squamosina to be connected in population from the Red Sea all the way South to Mozambique, as there are natural barriers which would prevent its planktonic larvae from riding currents to intermix between the two regions. When populations are separated by a barrier, the flow of genes between them is cut off and evolution begins to separate the populations from each other until they are separate species, a process called allopatric speciation.
A large specimen of T. elongatissima observed by iNaturalist user dawngoebbels off of Kenya
I figured that someday, researchers would collect tissue samples from these mystery clams to settle whether they were actually T. squamosina or something else. And this year, a team did just that, traveling along the coast of Mozambique, Madagascar, Kenya and other places, collecting samples of tissue to compare how all the different clams they saw were related in a family tree. They genetically sequenced these “clamples” and in the process, found that the mystery clams were a new cryptic species, which they called T. elongatissima!
Shells of T. elongatissima from the Fauvelot et al. 2020 paper
For comparison, a shell of T. squamosina collected off of Sinai, Egypt. You can see why they’re easy to mix up!
T. elongatissima closely resembles T. squamosina, and they are sister species on the bivalve family tree. It’s hard to tell them apart without training. Even a professional would probably mix some of them up if they were all placed sitting next to each other. The major differences appear to relate to shell shape, with T. elongatissima having a less symmetrical shell than T. squamosina, and a bigger opening at the rear hinge for a foot to poke through. The symmetrical shell and closing of the foot opening may represent changes that T. squamosina took on to adapt to be able to sit freely on the bottom, rather than embedding in the coral like T. elongatissima seems to prefer. If you’ve read this far, you may be thinking “Who cares? A clam’s a clam and these look practically the same. Aren’t you just splitting clams at this point?” At the end of the day, a species is a man-made concept; an organizing tool for use by us humans. Species are the characters in our reconstruction of the history of the world. What can we learn about the world by having identified this species T. elongatissima?
A giant clam family tree! Notice T. squamosina and T. elongatissima right next to each other.
The researchers behind the new paper discuss that based on statistical analyses of the genetic differences between the species, the most recent common ancestor for T. elongatissima and T. squamosina probably lived more than 1.4 million years ago! Some researchers have previously suggested that T. squamosina probably began its development as a separate species due to geographic isolation by low sea level, caused by repeated glaciations. With so much water trapped as ice on land during this period, the narrow Strait of Bab al Mandab, currently the gateway to the Red Sea, became a land barrier as sea level fell (kind of like opposite of the Bering Sea land bridge that formed allowing humans to migrate to the Americas). Ancestral clams trapped on the Northern end of this barrier were proposed to have evolved to become the rare T. squamosina.
This has occurred with a variety of species that became Red Sea endemics (meaning they are unique species that evolved in the Red Sea and are found nowhere else), including a unique crown of thorns starfish. The issue is that during this time of low sea level, the Red Sea went through periods where it was a rather unfriendly place for clams to live. All sorts of creatures went extinct in the period when the sea was repeatedly cut off, because the water became extremely salty, along with other unfriendly changes. So it’s unlikely T. squamosina would be present for us to see today if it only lived in the Red Sea throughout the entire length of time.
A map from Fauvelot et al. 2020 showing the distributions of different giant clams the researchers identified along the coasts of Africa and the Red Sea. Notice the bright red dots representing T. squamosina, only found in the Red Sea, while green dots represent T. elongatissima. Notice how the currents (arrows) seem to meet and then go offshore from Kenya. More on that in the next paragraph.
The researchers of this new paper propose that T. squamosina was more likely to have initially branched off due to the barrier of the Horn of Africa. The seas off of Kenya and Somalia harbor a meeting of southward and northward currents which then group and head offshore, away from the reefs that giant clam larvae are trying to get to. So any tiny floating planktonic clam larvae would experience a strong “headwind” preventing them from crossing that point. It would also mean that during times that the Red Sea was not a happy place to be a clam, T. squamosina may have found refuge on the coasts of places like Eritrea, Oman and possibly even as far as Pakistan. During times when sea levels rose and Red Sea conditions became friendlier, it recolonized the area.
As far as we know, the Red Sea is the only place T. squamosina is now found, but it may well be present elsewhere like Yemen or Oman. If T. squamosina was found in other regions, it would be tremendously important for its conservation. Right now, the species is thought to be extremely rare, with a very small native range. If it inhabited a broader area, that would mean more reservoirs of genetic diversity. This would reduce the odds that it will go extinct as reefs are put under stress from climate change, pollution and overharvesting. To survive as a species, it helps to not put all your eggs in one basket. If you’re only found in one small place, it increases the chances that a disaster (like climate change) will wipe you out.
The only way we will know for sure is to visit reefs in understudied places like Yemen, Oman, Pakistan, Eritrea and Somalia, to understand the richness of the giant clams present. These areas are understudied for various reasons: lack of research funding for non-Western researchers, lack of interest from the scientific community too focused on familiar places, and geopolitical situations that make it difficult to conduct research. But I hope someday to collaborate with people in these countries to better understand the giant clams present in such understudied regions of the globe. It is virtually certain that there are more species of giant clams, both alive and as fossils, waiting to be discovered.
Is eating these a gamble? Science can help improve our odds!
I am often asked if I eat clams. The answer is yes: while I love to observe live clams and appreciate their abilities, I will eat a good clam chowder or plate of grilled scallops if presented with the chance. While I’m generally not a fan of super fishy-tasting foods, I eat bivalves with a clear conscience because farmed mollusks represent a super sustainable way to get protein! However, as many of us have learned the hard way, shellfish can sometimes produce unwanted results later after the meal, if the animals are contaminated with food poison. Eating such “bad” clams can produce a spectrum of food poisoning symptoms ranging from vomiting and diarrhea to memory loss to even paralysis and death.
Humans have known the hazards of eating shellfish for a very long time. It has been suggested that the ban on shellfish present in kosher and halal dietary rules arose as a preventative measure to protect from food poisoning (though eating fish, land animals and even vegetables can poison people in numerous ways as well). Studies of oysters have determined that ancient peoples of modern day Georgia from 5000 years before present selected their season of harvest based partially on knowledge of the seasons when such poisoning was most prevalent in their area.
How and why does this happen, and what can we do to prevent it? It’s a billion-dollar question, because when flare-ups of shellfish food poisoning happen, they are hugely costly to fishermen and the food industry, costing millions of dollars a year in lost business when fisheries are forced to shut down and products are recalled. Such events are increasing in frequency and severity. Which makes it all the more strange that these shellfish poisoning events are not the fault of the bivalves per se, but rather what they’re eating.
Note: people generally get annoyed when you start to point out the body parts of the oyster they’re about to swallow whole. Source
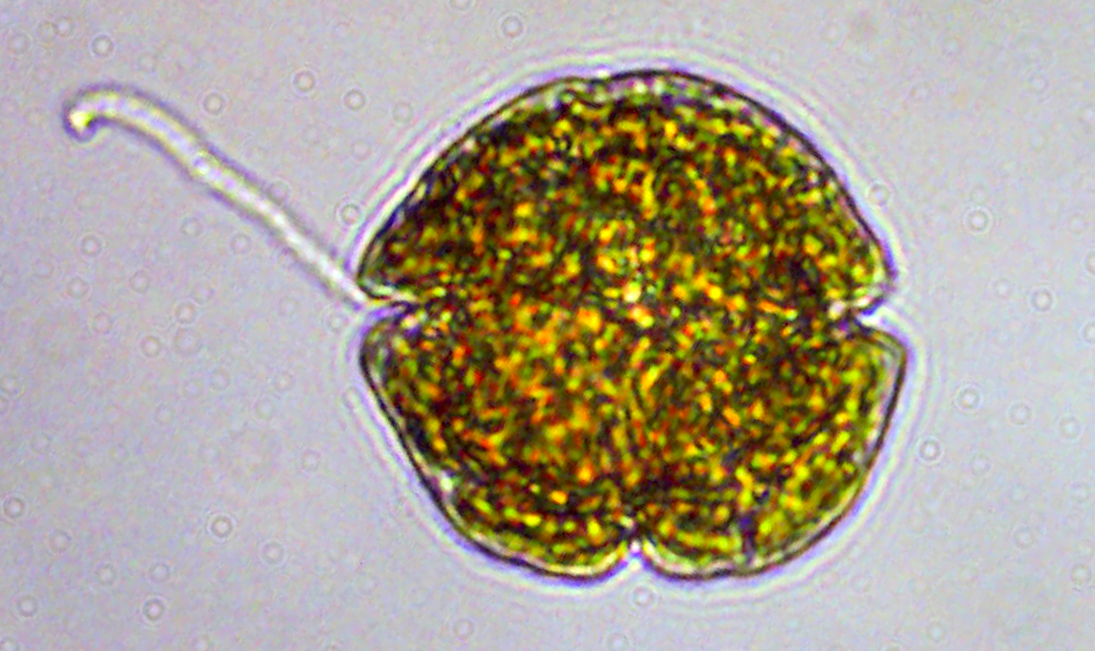
Almost all bivalves are filter-feeders, using their gills to gather small passing food particles, which they then either ingest or discard based on the quality of the food item. Clams are cows crossed with Brita filters, and for many species of clams which we eat, the reason they do all this filtration is to find phytoplankton food. Phytoplankton are microscopic algae suspended by ocean currents that make their living from photosynthesis. They are a hugely plentiful and high-quality food item, making up a huge amount of the biomass available in the ocean. Like plant-life on land, phytoplankton are highly seasonal in their appearance, rising and falling in abundance in periodic “bloom” events.
Aerial view of a red tide off the Texas coast. Source: NOAA
But as Spongebob Squarepants taught us, plankton are not always peaceful. Many types of algae produce toxic compounds which may be integrated into the body parts of bivalves that eat them. Scientists call the blooms of algae which produce toxins “Harmful Algal Blooms” (HABs), and such events are growing in frequency and cause huge harm to marine life and sicken thousands of people per year. There are many algae species which cause HABs all around the world, sometimes visible as “red tides,” but not always. When HABs occur, they can lead to mass deaths of higher animals in the food chain that feed on clams such as marine mammals and seabirds. In fact, HABs are at their most dangerous to humans when they catch us by surprise.
Who me? I’d never!
Microscope view of the toxic dinoflagellate Karenia. Source: NOAA
When humans eat bivalves which have been dosed with such marine toxins, many types of poisoning can occur. Brevetoxin is produced by a type of dinoflagellate phytoplankton Karenia as well as other species, and when humans are exposed, we can suffer from Neurotoxic Shellfish Poisoning, which causes vomiting, diarrhea and even neurological effects like slurred speech. Saxitoxin is produced by a variety of plankton species including dinoflagellates and freshwater cyanobacteria. When ingested in clams (such as the butter clam Saxidomus which gave it its name), fish or other animals, it can cause Paralytic Shellfish Poisoning, a sometimes fatal syndrome which shuts down nerve signaling, leading to temporary paralysis.
So we know it’s bad for humans to ingest these toxins. What is it doing to the clams? Oddly enough, some types of toxins like saxitoxin are not that harmful to the clams or other plankton eating animals, allowing them to accumulate huge amounts in their bodies with little ill effect. Its presence does not seem to influence their feeding behavior much, or their growth after exposure. Its status as a neurotoxin in mammals might be a total chemical and evolutionary coincidence, as researchers suggest that it may actually serve as a signal in some part of the algae’s mating cycle. This also may be the case for brevetoxin, which appears to be produced when Karenia is under environmental stress. But there is not much agreement in the HAB and aquaculture research fields, because there are many types of algae, which may produce their toxins for many reasons, and it is very hard for us to zoom in to the scale of the microbe and out to the scale of the ecosystem at the same time, to find any kind of universal evolutionary role of these toxins. Some researchers insist that some bivalves are influenced negatively by brevetoxin, but only at the juvenile stage during major bloom events. The effects of the toxin may only influence certain species, or only become significant if the toxin reaches the digestive tract of the bivalve. Overall, research into impact of HABs on clams is still a topic of active research, and the idea that the microbes produce these toxins to defend against bivalve predators is definitely not a slam-dunk, easily proven hypothesis. While some clams are negatively affected by the toxins, it is not consistently observed across species in a open-and-shut way, and it can be a subtle effect to observe and quantify scientifically.
Karenia to mammals: Oops!
The more I read about this stuff, the more shocked I am at the incredible complexity of marine algae and their toxins. I only started reading about them trying how to to understand how they influence bivalves. I was hoping to find some evidence of their effects on bivalve growth that I could apply back in time in fossil shells to understand the historical occurrence of HAB events. It’s important to understand HABs because they hurt people, cost our society a lot of money and if we understand how to avoid them, we can help minimize such impacts in the future as HABs continue to become more common.
Hi colleagues! Several weeks ago, I was supposed to present a talk at GSA’s annual meeting in Phoenix at the session “Advances in Ocean and Climate Reconstructions from Environmental Proxies”, but I shattered my wrist in a scooter accident the night before and was in emergency surgery during my talk time. So instead I’ve uploaded my talk with voice-over to Youtube! The whole video is about 15 minutes. You can view it above. Feel free to comment on this post or email me if you have questions!
This work is currently in the last stretch of drafting before submission, but I also discuss some ongoing research and am always open if you have your own ideas for collaborations!
Correction: we are working with geophysicists to understand the shell transport mechanism.
These are the references mentioned at the end:
Crnčević, Marija, Melita Peharda, Daria Ezgeta-Balić, and Marijana Pećarević. “Reproductive cycle of Glycymeris nummaria (Linnaeus, 1758)(Mollusca: Bivalvia) from Mali Ston Bay, Adriatic Sea, Croatia.” Scientia Marina 77, no. 2 (2013): 293.
Grossman, Ethan L., and Teh-Lung Ku. 1986. “Oxygen and Carbon Isotope Fractionation in Biogenic Aragonite: Temperature Effects.” Chemical Geology: Isotope Geoscience Section 59: 59–74.
Gutierrez-Mas, J. M. 2011. “Glycymeris Shell Accumulations as Indicators of Recent Sea-Level Changes and High-Energy Events in Cadiz Bay (SW Spain).” Estuarine, Coastal and Shelf Science 92 (4): 546–54.
Jones, Douglas S., and Irvy R. Quitmyer. 1996. “Marking Time with Bivalve Shells: Oxygen Isotopes and Season of Annual Increment Formation.” PALAIOS 11 (4): 340–46.
Mienis, Henk, R. Zaslow, and D.E. Mayer. 2006. “Glycymeris in the Levant Sea. 1. Finds of Recent Glycymeris insubrica in the South East Corner of the Mediterranean.” Triton 13 (March): 5–9.
Najdek, Mirjana, Daria Ezgeta-Balić, Maria Blažina, Marija Crnčević, and Melita Peharda. 2016. “Potential Food Sources of Glycymeris nummaria (Mollusca: Bivalvia) during the Annual Cycle Indicated by Fatty Acid Analysis.” Scientia Marina 80 (1): 123–29.
Peharda, Melita, Marija Crnčević, Ivana Bušelić, Chris A. Richardson, and Daria Ezgeta-Balić. 2012. “Growth and Longevity of Glycymeris nummaria (Linnaeus, 1758) from the Eastern Adriatic, Croatia.” Journal of Shellfish Research 31 (4): 947–51.
Reinhardt, Eduard G, Beverly N Goodman, Joe I Boyce, Gloria Lopez, Peter van Hengstum, W Jack Rink, Yossi Mart, and Avner Raban. 2006. “The Tsunami of 13 December AD 115 and the Destruction of Herod the Great’s Harbor at Caesarea Maritima, Israel.” Geology 34 (12): 1061–64.
Royer, Clémence, Julien Thébault, Laurent Chauvaud, and Frédéric Olivier. 2013. “Structural Analysis and Paleoenvironmental Potential of Dog Cockle Shells (Glycymeris glycymeris) in Brittany, Northwest France.” Palaeogeography, Palaeoclimatology, Palaeoecology 373: 123–32.
Sivan, D., M. Potasman, A. Almogi-Labin, D. E. Bar-Yosef Mayer, E. Spanier, and E. Boaretto. 2006. “The Glycymeris Query along the Coast and Shallow Shelf of Israel, Southeast Mediterranean.” Palaeogeography, Palaeoclimatology, Palaeoecology 233 (1): 134–48.
When I mention to people that I study bivalves, I can sometimes sense from their facial expressions that they are secretly asking “why?” While clams are perfectly content to keep doing what they’re doing without being thanked, I think it’s important to enumerate all of the ways they make our world more livable and functional.
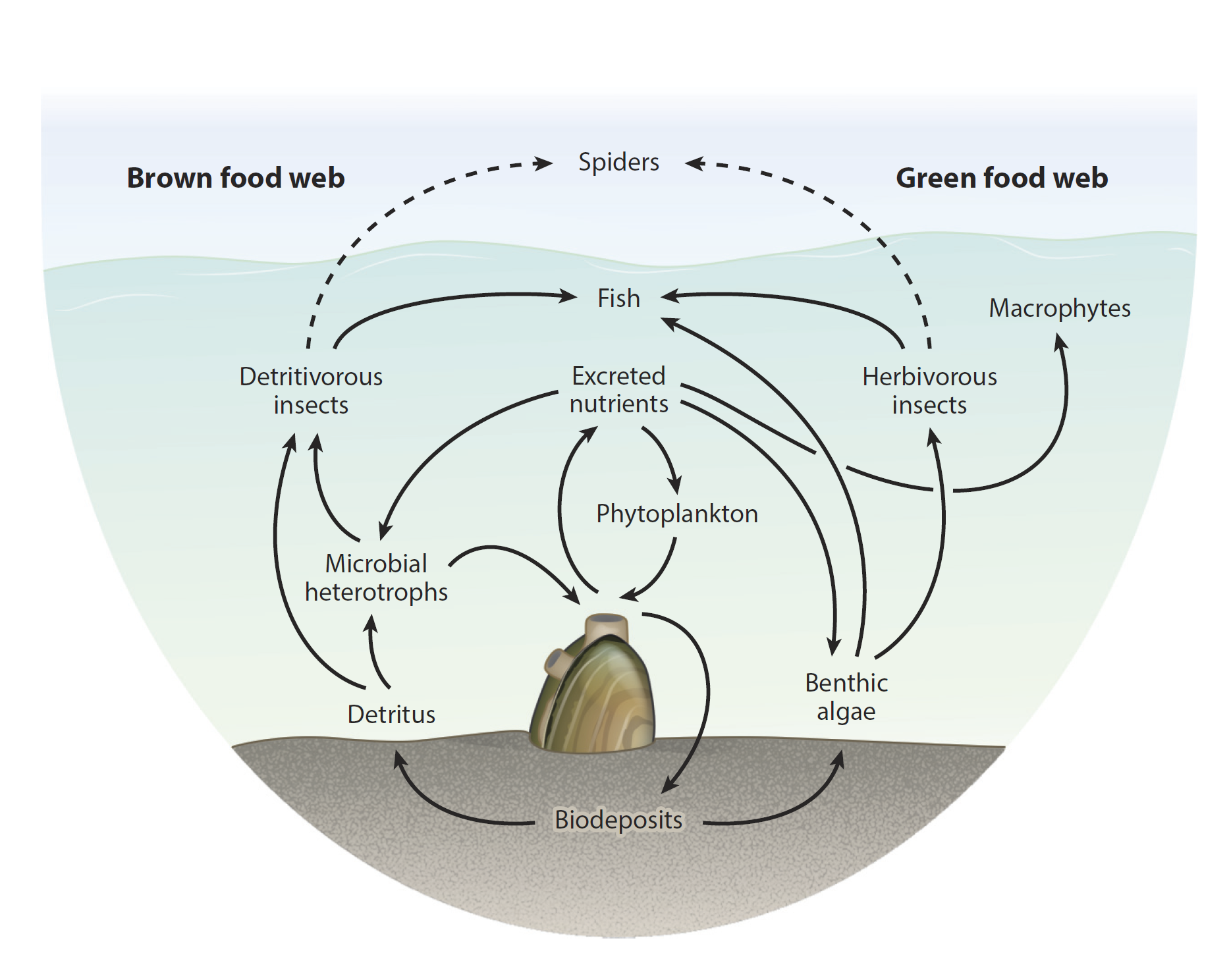
Various roles that freshwater mussels can play in their local food webs (Source: Vaughn and Hoellein, 2018)
Bivalves are ecosystem engineers. While they may seem rather stationary and not up to much at any particular time, they are actually always working to actively maintain their habitat. The majority of clams are filter-feeders, meaning that they use their gills to gather particles from the water column for food. Some of these particles are ingested as food and later pooped out. Some inedible particles are discarded immediately by the clam as “pseudofeces”. Both mechanisms serve as a bridge between the water column and the benthos (the sediment at the bottom). In this way, clams are engines that take carbon fixed by algae floating in the water and transfer that material to be stored in the sediment. Their bodies also act as nutrition to feed all sorts of animals higher on the food chain like sea stars, lobsters, seabirds, sea otters and humans that depend on bivalves as food. They are literally sucking up the primary productivity (algae) to be used by the rest of the food chain.
The filtration rate of oysters. Graphic from The Nature Conservancy
Different clam species vary in their precise filtration rate (how fast they can inhale and exhale water, filtering the particles within), but it is prodigious. Some freshwater mussels, for example, can pick-through 1-2 liters of water per hour for every gram of their own flesh. Since these individual bivalves can weigh over 100 g, they are capable of picking the food out of an immense quantity of water. In doing so, bivalves help improve the clarity of the water column, allowing more sunlight to reach deeper into the water body (the photic zone), providing more energy for additional photosynthesis to occur. While there are examples where invasive bivalves such as zebra or quagga mussels take this phenomenon too far, in well-functioning ecosystems, the filtration activity of clams helps improve the productivity of the community.
An oyster reef. Source: The Nature Conservancy
Bivalves help make sediment through their filtration of material from the water column, and they also engineer and manipulate the sediment directly. Some bivalves, like oysters, are able to make huge mounds of dirt that serve as habitat for all sorts of life, increasing the diversity of the community. They do so both by excreting sediment, and also by passively trapping it between the shells of neighboring oysters (“baffling”). By doing so, they reduce rates of coastal erosion and increase the biodiversity of wetlands. For this reason, New York and other communities plan to seed oyster reefs to help fight sea level rise and reduce the threat of storm surges like the one that occurred during Superstorm Sandy.
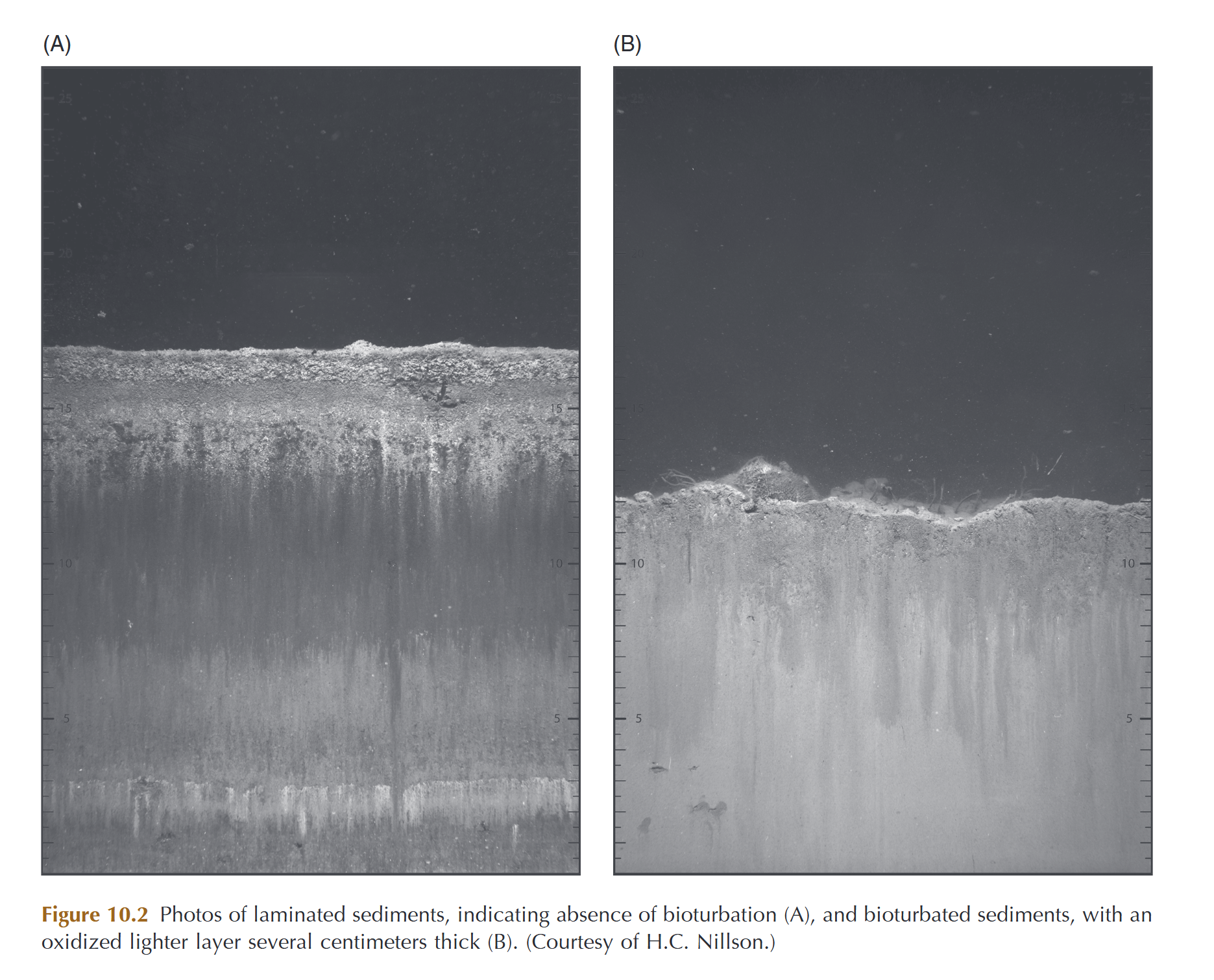
Comparison of sediments without bioturbation by digging animals, and with. Notice how the non-bioturbated sediment is layered and darkened due to activity by anaerobic bacteria, while the well-oxygenated, mixed sediment is light all the way through. From Norkko and Shumway, 2011
Other “infaunal” bivalves (burrowers) help to aerate the sediment through their tunneling, bringing oxygen deep under the surface of the dirt. This mixing of the sediment (also called bioturbation) ensures that nutrition from deep under the sediment surface is again made available for other organisms. Some bivalves can bore into coral reefs or solid rock, creating burrows which serve as habitat for other animals and can free up minerals for use by the surrounding ecosystem. Helpful shipworms assist in eating wood, assisting in returning nutrients stored in that tissue to the ecosystem as well.
Enormous grouping of giant clams in a lagoon in French Polynesia. From Gilbert et al., 2005
Bivalves of course are also famous for their shells, and this activity also provides habitat to sponges, snails, barnacles and many other encrusting organisms specially adapted to live on bivalve shells and found nowhere else. Giant clams are the most legendary “hypercalcifiers,” and in some regions like New Caledonia can rival reef-building corals in terms of biomass. In areas where soft-bottoms dominate, bivalves like hammer oysters, adapted to “rafting” on the quicksand-like surface of the soft sediment, can assist by providing a platform for other animals to take refuge. In the deep sea, bathymodiolid mussels and other chemosymbiotic bivalves can feed directly on the methane and sulfur emitted from hot vents or cold seeps with the help of symbiotic bacteria, creating dense reefs which provide food and habitat for all sorts of life. Even once the clams die, their shells can continue to serve as homes for other creatures.
Crabs feeding on Bathymodiolus in the deep sea (NOAA)
The shells of clams provide great scientific value in understanding our world. Much like tree rings serve as a record of environment thousands of years into the past, growth rings in clam shells serve as a diary of the animal’s life. These rings can be yearly, lunar, tidal or even daily in rhythm, with each ring serving as a page in the diary. The chemistry of those “pages” can be analyzed to figure out the temperature the clam experienced, what it ate, whether it suffered from pollution, and even the frequency of storms! The study of rings in the hard parts of animals is called sclerochronology, and it’s what first drew me to study bivalves. I was so fascinated by the idea that our beaches are covered with high-resolution records of the ocean environment, waiting to be cut open and read.
This giant clam shell recorded an interruption in the animal’s daily growth caused by a typhoon! From Komagoe et al., 2018
While they don’t owe us anything, clams provide a lot of value to humans as well, serving as a sustainable and productive source of food. Humans have been farming bivalves for thousands of years, as evidenced by “oyster gardens” and shell middens which can be found all over the world. Particularly in seasons when food is scarce on land, native peoples could survive by taking advantage of the wealth of the sea, and bivalves are one of the most plentiful and accessible marine food sources available. But they aren’t just the past of our food; they may be part of the future. Bivalves are one of the most sustainable sources of meat known, requiring very little additional food to farm and actively cleaning the environment in the process. Mussels grown out on a rope farm are an easy investment, growing quickly and with very little required energy expenditure. Someday, giant clams may provide the first carbon-neutral meat source, as they gain their food from symbiotic algae within their flesh. I have never eaten one, but I’ve heard they’re delicious.
A shell midden in Argentina. Photo from Mikel Zubimendi, Wikipedia
Mussels being farmed on ropes
Clams are heroes we didn’t know we needed and maybe don’t deserve. They ask for nothing from us, but provide vast services which we take for granted. So the next time you see an inconspicuous airhole in the sand, thank the clam that could be deep below for aerating the sediment. The shell of that long-dead mussel at your feet may have fed a sea star, and now is a home for barnacles and many other creatures. While that mussel was alive, it sucked in algae to improve water quality on our beaches. And the sand itself may contain countless fragments of even more ancient shells. Clams silently serve as an important cog in the vast machine that makes our oceans, rivers and lakes such amazing places to be. Thank you clams!
To us active, dynamic mammals, the humble clam can appear positively…inanimate. Their nervous system is decentralized relative to ours, lacking any sort of brain, and to the untrained eye, it can appear that their only discernible reaction to the outside world is opening or closing. Open = happy, closed = not happy; end of story, right? Some vegans even argue that the clams are so nonsentient that it is okay to eat them and think of them as having no more agency than a vegetable!
You might already have predicted I intend to tell you about just how animate and sentient clams can be. But let’s start out by describing the nuts and bolts of their nervous system. As with many invertebrates, their nervous system is distributed throughout their body as a system of ganglia. Ganglia are clumps of nerve cells which may have local specialization, and transmit messages within neurons using electrical potentials. At the connection between cells (called a synapse), neurotransmitters are used to pass signals to the next cell. Researchers have found that bivalves use “histamine‐, octopamine‐, gamma‐aminobutyric acid‐ (GABA)…like immunoreactivity” in their central and peripheral nervous systems, much like us vertebrates do, and other studies have even found that the response to serotonin and dopamine is localized in nervous tissue linked to different organ systems.
Nerve cells (bright green) highlighted in a larval oyster with fluorescent dye (from Yurchenko et al 2018)
These systems of chemical nerve transmission are truly ancient, likely dating back to the formation of complex animal body plans in the earliest Cambrian. Researchers have great interest in studying these nervous and hormonal signaling systems in mollusks because they can shed light on the relative flexibility and limitations of these systems throughout the animal tree of life. Characterizing these systems can also allow us to understand the mechanisms that bivalves and other animals use to react to environmental stimuli.
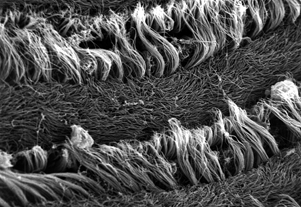
Electron microscope view of gill cilia, zoomed in 1000x (from Dan Hornbach)
Like humans, bivalves spend a lot of time and effort eating. Most bivalves eat by filtering food from passing water with tiny cilia on their gills. These cilia work to capture food particles and also act as a miniature rowing team moving water along the gill surface. The bivalve needs a way to control this ciliar activity, and researchers found they could directly control the speed at which oysters move their cilia by dosing them with serotonin and dopamine, which respectively increased and decreased activity.
Bivalves also work very hard to make babies. Most bivalves reproduce by releasing sperm and eggs to fertilize externally in the water column. To maximize their chances to find a mate, they typically save up their reproductive cells in gonads for multiple months and release them in a coordinated mass spawning event. It appears that this process is controlled by hormonal releases of dopamine and serotonin. Researchers have determined that serotonin concentrations vary through the year, with mussels in New England using it to regulate a seasonal cycle of feeding in summer, followed storing of that energy for winter. During the winter when food is less available, they use that stored energy to bulk up their gonads in time for reproductive release in spring months, when their larvae have plentiful access to food and oxygen, ensuring them the best chance of survival. In recent decades, aquaculturists have learned to use serotonin injections to induce spawning in cultured clams, to ensure they will have a harvest ready at a certain time of year.
So bivalves are very sensitive to the seasons. How about shorter term sources of excitement? You might have observed this yourself through the clam’s most iconic activity: opening and closing its shell. Clams close their shells with powerful adductor muscles which pull the two valves together. A springy ligament at the hinge pulls the shell open when the muscles relax. Just like us, the clam needs to use nerve cells to signal the muscle to do its thing. In addition, two different sets of ganglia act to control the foot that some bivalves can extend to dig into sand, with one ganglion acting to extend the foot and the other causing it to contract. While clams don’t have a centralized brain with specialized regions for different uses like we have, this represents a sort of specialization of neural systems with a similar result.
This iconic gif is often shared along with the claim it shows a clam “licking” salt. It is actually using its foot to search for a place to dig. The salt was not needed.
When a certain neuron is used repeatedly, it can form a cellular memory allowing the organism to acclamate (ugh sorry) and moderate its response to a particular stimulus over time. Giant clams, for example, close their shells when their simple eyes detect a shadow overhead. This behavior can protect them from predation. When I conducted some of my PhD research, sampling body fluid of aquarium and wild giant clams with a syringe, I noticed that captive clams didn’t close up in response to my shadow overhead, while wild clams required me to sneak up and wedge their shells open with a wooden block to do my work. I suspected that after exposure to frequent feedings and water changes by aquarists, the clam had “learned” that there was no reason to expend energy closing its shell. Meanwhile, in the process of proving that our sampling technique was not harmful to the animal, I discovered that clams which detected my shadow would quickly reopen within seconds when I hid from them, while those that were stuck by a syringe would stay closed for minutes before opening and beginning to feed again. Makes sense!
Other researchers noticed this phenomenon as well. One group found that giant clams repeatedly exposed to shadows of different sizes, shell tapping and even directly touching its soft tissue began to habituate (become accustomed) to the stress, opening more quickly and staying open longer each time the stimulus occurred. Even more interestingly, they did not transfer that habituation between stress types; for example, the clams that saw a shadow again and again would still react strongly to a different stress like tapping its shell. This suggests the animal can distinguish between different threats along a spectrum of seriousness, with touching of tissue (similar to a fish pecking at its flesh) being the most serious threat with the most dramatic response.
Another study determined that larger giant clams stayed closed longer than smaller ones in response to the same threat. They proposed this was related to the greater risk large clams face as they have more tissue area vulnerable to attack. While the clams might not have made a “conscious” decision in the way we do as thinking creatures, they were able to place their individual risk in context and vary their response. This ability to tailor a response to different risk levels is a sign of surprisingly complex neurology at work.
Close up of the eyes of a scallop. Each is a tiny crystalline parabolic mirror (photo by Matthew Krummins on Wikipedia)
Scallops show some of the most complex bivalve behaviors. This relates back to their unique adaptations, including simple eyes that can resolve shapes and the ability to swim away from danger. Scallops have been found to discern between predator types by sight alone, closing or swimming away in response to crustaceans but not sea stars. When swimming, they are capable of using this vision to navigate to places where they can hide, such as seagrass beds. They even use their vision to gather food: in one of my favorite experiments, researchers showed scallops a video of food particles, and they opened their shells and started filter feeding in response, kind of like me salivating when I watch the Food Channel! It would be very interesting to compare the behavior of scallops in marine protected areas to those that can be freely harvested. Do they vary their behavior in response?
I hope I’ve made clear that while clams are not exactly intellectual powerhouses, their behavior is much more complicated than simply sucking up water and opening or closing their shells. Like us, they inhabit a complex environment that requires a multitude of responses. Their nervous systems have evolved to allow them to survive and adopt nuanced behaviors which they can vary on the fly, and which us “higher” animals are only just beginning to comprehend.
Scallops spooked by divers’ lights and fleeing en masse to filter somewhere else
The ocean is a place of constant dynamic movement. Fish use their fins to push water away from themselves, and because every action has an equal and opposite reaction, they therefore move forward. Some cephalopods use jet propulsion, constricting their mantle cavity to push water out through siphons, allowing them to jet forward like a deflating balloon. And other life forms sail the seas on constantly moving currents , indirectly harnessing the power of the sun and earth.
Bivalves are a fairly sedentary bunch by comparison. While most bivalves have a planktonic larval form, when they settle they are constrained to a fairly small area within which they can burrow or scramble around with their muscular feet.
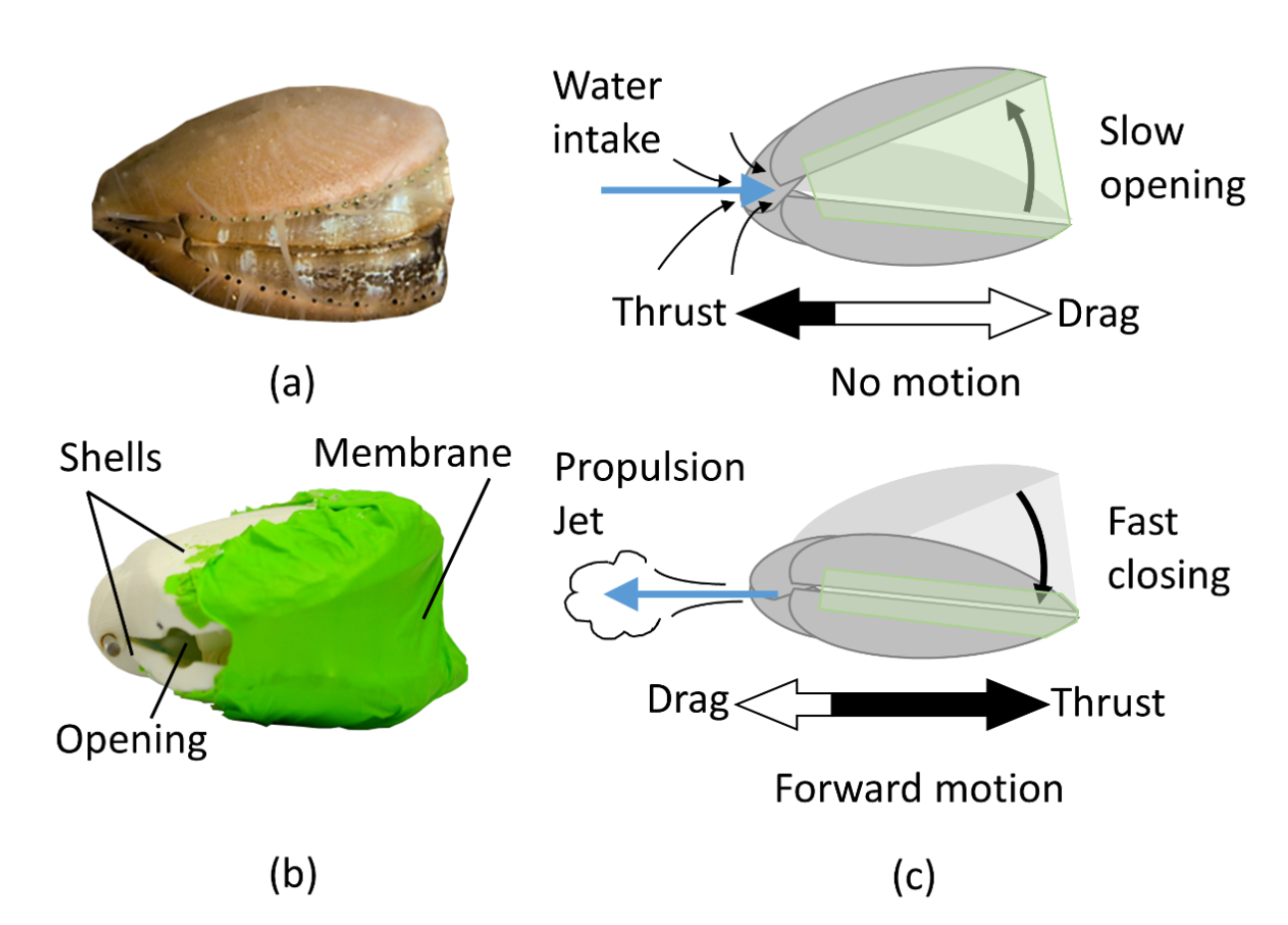
But some bivalves have evolved to move at a quicker rate. The most famous swimming bivalves are the scallops, which have evolved to use jet propulsion, similar to their very distantly related cephalopod relatives. But unlike the cephalopods, scallops evolved to use their hinged shells to aid this process!
Notice the expelled water disturbing the sediment below the scallop as it “claps” its way forward!
Many filter-feeding bivalves use their shell valves as a biological bellows to pull in water for the purposes of sucking in food, or even to aid in digging, but scallops have developed another use for this activity, to enable propulsion. Scallops draw in water by opening their valves to create a vacuum which draws in water to their sealed mantle cavity. They then rapidly close their valves using their strong adductor muscles to pull them together, which pushes the water back through vents in the rear hinge area, propelling the scallop forward.
Don’t panic if a scallop swims toward you. They can see, but not super well. This one is just confused.
Using this strategy, scallops can evade predators and distribute themselves to new feeding sites. It’s a surprisingly effective swimming technique, with the queen scallop able to move 37 cm/second, or over five body lengths per second! Michael Phelps would have to swim at nearly 35 km/h to match that relative speed (his actual highest speed is around 1/3 of that). I’m sure sustaining that speed would be tiring for Mr. Phelps, though, and it’s the same for scallops, only using their swimming for short-distance swims.
A recent paper from a team in Switzerland just came out describing an effort to engineer a robot which imitates the scallop’s elegant and simple swimming method. The resulting totally adorable “RoboScallop” closely imitates the design of a scallop, using a pair of hinged valves with rear openings to allow the movement of water backward. The internal cavity is sealed by a rubber membrane draped across the front so that all water is forced through these rear vents when the Roboscallop snaps shut.
Diagram from the Roboscallop paper (from Robertson et al. 2019)
As seen in the diagram above, the rhythm and relative velocity of opening vs closing is important to make sure the RoboScallop actually moves forward. If the scallop opened as quickly as it closed, it would just rock back in forth. It instead opens slowly so that it does not draw itself backward at the same rate that it can push itself forward. The researchers had to do quite a bit of calibration to get these rates right (equating to about 1.4 “claps” per second), but once they did, they ended up with a RoboScallop that can generate about the same force of forward movement (1 Newton) as a real scallop (1.15 Newtons), and similar rates of speed.
This paper really fascinated me because it is merely the latest in a long line of successful engineering projects imitating the ingenuity of evolution. Other marine robots have been made which emulate the locomotion of fish, manta rays, sea snakes and other forms of swimming. And now we have a clam! Let me know when I can buy one to play with in my pool.
If you’ve read any of my posts, you should realize by now that clams are pretty weird. Some catch live prey. Some have algae in their bodies that they “farm” for food. Some can bore into hard rock. Some sail the seas on rafts of kelp. Clams live in a competitive world and have had hundreds of millions of years of time to evolve to try out all sorts of weird, unlikely ways of life.
The thick shelled river mussel (Unio crassus) is known from many rivers and streams of Central Europe. As this is a very well-studied region of the world, many generations of academics have noted an unusual, seemingly inexplicable behavior undertaken by these mussels at certain times of year.
Using its muscular foot, U. crassus pulls itself to the edges of the streams and rivers it lives in until it is partially exposed to air. It orients itself at a right angle with the surface of the stream with its siphons (two little snorkels coming out of the shell) facing out towards the water. Like all bivalves, U. crassus can act as a bellows by opening and closing its shell to pull in and push out water through those siphons. It has one siphon above the water and one below, and it proceeds to suck in water and spray it into the center of the stream using the power of its suction. The water can travel over a meter away and they continue this spurting about once a minute, sometimes for hours.
Needless to say, this is a very strange and unlikely behavior to observe in a mussel. It is exposing itself to potential dessication or suffocation from exposure to air. It is vulnerable to predation from terrestrial mammals and birds. There has to be a very powerful benefit from this behavior to outweigh those risks. And why squirt water into the air?
Some researchers proposed that the mussels were traveling to shore to harvest from the more plentiful food particles deposited there. But why would they face their siphons away from the shore then? Other workers suggested that it was a way to reduce heat stress through evaporation, though that also seems unlikely, considering the water is warmest in the shallows. The question persisted for decades in the minds of curious malacologists.
In 2005, Heinrich Vicentini of the Swiss Bureau for Inland Fisheries and Freshwater Ecology decided to try settle the question of why these mussels spurt. He observed several dozen of the mussels crawl to the edge of the water and diligently begin squirting into the streams. In the name of science, he put himself in the path of these squirts, caught the water and used a hand lens to observe that the squirted water was full of mussel larvae (glochidia).
U. crassus falls in the order Unionida, a group of freshwater mussels distinguished by a very unusual method of reproduction. They are parasites! Because they can’t swim well enough to colonize upstream against the current, they need to rely on fish to hitch a ride. Some have evolved elaborate lures to convince fish to take a bite, then allowing them to release their larvae, which attach to the fish’s gills like binder clips and ride all the way upstream. Once they have reached their destination, they detach and grow up into more conventional burrowing mussels. It’s a weird, creepy and wonderfully brilliant strategy that enabled the mussels to invade the inland rivers which would otherwise be inaccessible to them.
Loach (type of freshwater fish) gills with unionid larvae attached (UCforLife)
The mussels appear to be spurting out not only water, but their babies. They gain a couple of advantages from this. For one, their larvae can distribute further than would be possible from the bottom of the creek. Instead, they are released at the center of the surface of the stream, where they can be carried for a much longer distance by the current before they settle at the bottom. In addition, the splash of water on the surface may mimic the behavior of insects and other fish food falling in the water. A curious minnow might venture to investigate the source of the splash, where it would promptly breathe in a cloud of larvae that get stuck on its gills. A pretty rude surprise, but a brilliant trick to give the baby mussels the best chance of surviving.
So again, clams prove themselves to be far more clever and interesting than they might initially seem. U. crassus and other members of the Unionida are an ancient and globally distributed lineage which have evolved all sorts of weird and wonderful ways to maintain their river lifestyle. Unfortunately, rivers are some of the most widely damaged environments in the world. A majority of freshwater mussel species worldwide including U. crassus are endangered by habitat loss, overharvesting and pollution. But more research into their unusual biology can help us understand ways we can enhance their conservation, with the hope of providing more habitat for them to recover populations in the future. New projects in Sweden and other countries aim to recover habitat for their larvae to settle along 300 km of rivers, and research the fish species which their larvae prefer to hitch a ride on. With more work, we can hopefully ensure that the streams of Europe will harbor little mini super-soakers for millennia to come.
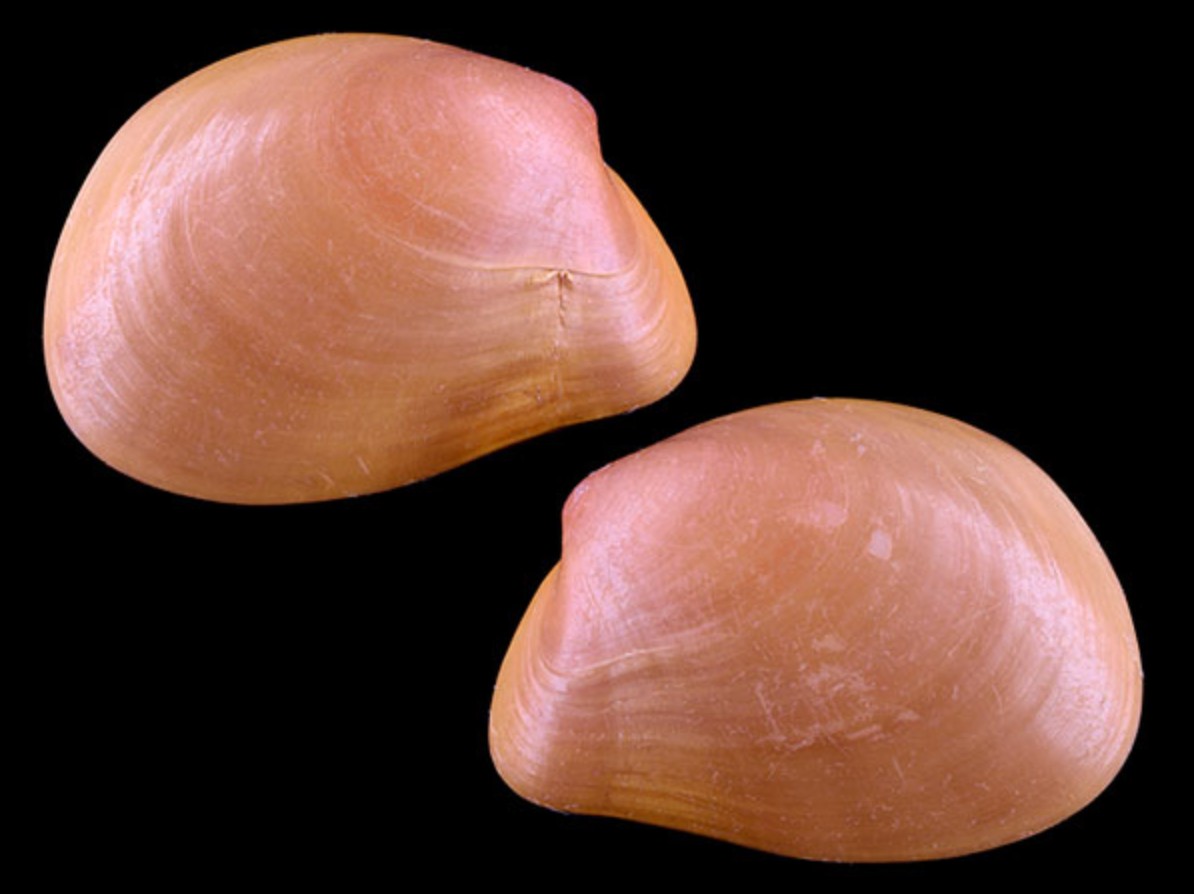
The streamlined shells of Gaimardia trapesina. Source: New Zealand MolluscaBivalves are not known as champion migrators. While scallops can swim and many types of bivalves can burrow, most bivalves are primarily sessile (non-moving on the ocean bottom). So for many bivalves, the primary method they use to colonize new territories is to release planktotrophic (“plankton-eating”) larvae, which can be carried to new places by currents and feed on other plankton surrounding them. Many bivalves have broad distributions because of their ability to hitchhike on ocean currents when they are microscopic. They don’t even pack a lunch, instead eating whatever other plankton is around them. But once they settle to grow, they are typically fixed in place.
Not all bivalves have a planktotrophic larval stage, though. Larvae of lecithotrophic bivalve species (“yolk-eaters”) have yolk-filled eggs which provide them with a package of nutrition to help them along to adulthood. Others are brooders, meaning that rather than releasing eggs and sperm into the water column to fertilize externally, they instead internally develop the embryos of their young to release to the local area when they are more fully developed. This strategy has some benefits. Brooders invest more energy into the success of their offspring and therefore may exhibit a higher survival rate than other bivalves that release their young as plankton to be carried by the sea-winds. This is analogous to the benefits that K-strategist vertebrate animals like elephants have compared to r-strategist mice: each baby is more work, and more risky, but is more likely to survive to carry your genes to the next generation.
Brooding is particularly useful at high latitudes, where the supply of phytoplankton that is the staple food of most planktrophic bivalve larvae is seasonal and may limit their ability to survive in large numbers. But most of these brooding bivalves stay comparatively local compared to their planktonic brethren. Their gene flow is lower on average as a result, with greater diversity in genetic makeup between populations of different regions. And generally, their species ranges are more constricted as a result of their limited ability to distribute themselves.
A bunch of G. trapesina attached to kelp. Notice the hitchhiking clams have in turn had hitchhiking barnacles attach to them. Freeloaders on freeloaders! Source: Eleonora Puccinelli
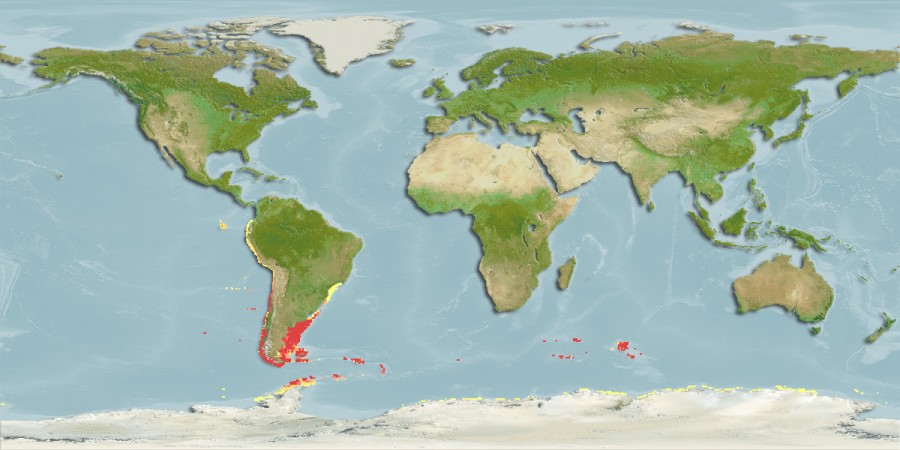
But some brooding bivalves have developed a tool to have it all: they nurture their young and colonize new territories by sailing the seas using kelp rafts. The clam Gaimardia trapesina has evolved to attach itself to giant kelp using long, stringy, elastic byssal threads and a sticky foot which helps it hold on for dear life. The kelp floats with the help of gas-filled pneumatocysts, and grows in the surge zone where it often is ripped apart or dislodged by the waves to be carried away by the tides and currents. This means that if the clam can persist through that wave-tossed interval to make it into the current, it can be carried far away. Though they are brooders, they are distributed across a broad circumpolar swathe of the Southern Ocean through the help of their their rafting ability. They nurture their embryos on specialized filaments in their bodies and release them to coat the surfaces of their small floating kelp worlds. The Southern Ocean is continuously swirling around the pole due to the dominance of the Antarctic Circumpolar Current, which serves as a constant conveyor belt transporting G. trapesina across the southern seas. So while G. trapesina live packed in on small rafts, they can travel to faraway coastlines using this skill.
The broad circumpolar distribution of G. trapesina. Source: Sealifebase
The biology of G. trapesina was described in greater detail in a recent paper from a team of South African researchers led by Dr. Eleonora Puccinelli, who found that the clams have evolved to not bite the hands (kelp blades?) that feed them. Tests of the isotopic composition of the clams’ tissue shows that most of their diet is made up of detritus (loose suspended particles of organic matter) rather than kelp. If the clams ate the kelp, they would be destroying their rafts, but they are gifted with a continuous supply of new food floating by as they sail from coast to coast across the Antarctic and South American shores. But they can’t be picky when they’re floating in the open sea, and instead eat whatever decaying matter they encounter.
Falkland Islands stamp featuring G. trapesina. Source.
The clams are small, around 1 cm in size, to reduce drag and allow for greater populations to share the same limited space of kelp. Their long, thin byssal threads regrow quickly if they are torn, which is a useful skill when their home is constantly being torn by waves and scavengers. Unlike other bivalves, their shells are thin and fragile and they do not really “clam up” their shells when handled. They prioritize most of their energy into reproduction and staying stuck to their rafts, and surrender to the predators that may eat them. There are many species that rely on G. trapesina as a food source at sea, particularly traveling seabirds, which descend to pick them off of kelp floating far from land. In that way, these sailing clams serve as an important piece of the food chain in the southernmost seas of our planet, providing an energy source for birds during their migrations to and from the shores of the Southern continents.
















































{kind=link}
{kind=link}
{kind=link}